We have been interested in testing for allelism among a collection of lethal defective kernel (dek) mutants. Because the mutants are lethal in the homozygous condition, it is necessary to cross heterozygous plants to test for allelism. Ideally each plant would be self-pollinated to determine whether or not the plant is heterozygous, and would also be cross-pollinated or would serve as a pollen source for cross-pollination. Since our stocks do not regularly bear two ears on each plant it is advantageous to be able to use a single ear to accomplish both a self-pollination and a cross-pollination.
Two procedures have been described for us where double-pollination (selfing and crossing) of a single ear is accomplished by pollinating a portion of the ear on one day and the remaining part of the ear the following day. The first of these methods used ears that are cut back in the usual manner. The silks visible on the first day of emergence are self-pollinated and the following day the additional silks that have emerged subsequent to the first pollination are cross-pollinated (M. G. Neuffer, pers. commun.). The second procedure involves cutting back an ear in the usual manner and self-pollinating the following morning. However, as soon as the pollination is completed a cut is made across the husks just above the top of the ear so as to sever the silks on one side of the ear. The following day the cut silks are brushed away and newly grown silks below the severed portions are then cross-pollinated (J. R. Laughnan, pers. commun.). Both of these procedures require returning to the female parent plant on a second day. In order to avoid this repeated visit, we have used a procedure where half of the silks on an ear are self-pollinated and half are cross-pollinated on the same day. This procedure equals or surpasses the results obtained with the above described procedures.
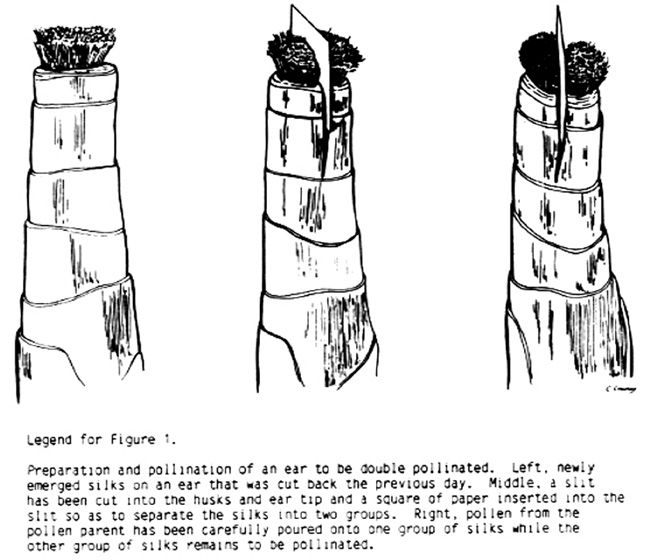
Double-pollination of a single ear is performed as follows: When the silks have completely emerged, they are trimmed back by cutting across the upper portion of the husks just above the tip of the ear in the usual fashion. Then a knife is used to slice downward a short distance into the rim of the husks and just into the ear tip to form a slit. A small paper square 3/4 inch x 3/4 inch (cut from a file card) is inserted into the slit to provide a separator for the silks that will subsequently emerge, dividing them into two equal parts. The slit in the husks and the ear tip holds the paper separator in a secure fashion (see Fig. 1).
Pollen from the cross donor is transferred to glassine bags to facilitate handling, the pollen parent is self-pollinated, and pollen is taken to the plants to be double-pollinated, where the pollen is carefully poured onto the silks on one side of the paper separator. The shoot bag is then replaced. After the cross-pollinations are completed, pollen is collected and used to accomplish self-pollination on the other side (Figure 1).
Discrimination of the cross-pollinated kernels from the self-pollinated kernels on a double-pollinated ear is aided by having the crossed kernels colored (anthocyanin) and the selfed kernels colorless. If the mutants to be tested are available in both colored and colorless stocks, ears can be produced with colored kernels on one half of the ear and colorless kernels on the other half. The colored kernels are the result of the cross-pollination and the colorless kernels are the result of the self-pollination.
An unambiguous test result is obtained when the self-pollinated ear of the pollen parent segregates for a dek mutant and the self-pollinated half of the female ear also segregates for a dek mutant. In such cases, the two mutants being tested are allelic if the cross-pollinated half of the ear of the female parent bears dek mutant kernels as well as normal appearing kernels, while the presence of only normal kernels on that half of the ear demonstrates that the two mutants are not allelic (see Figure 2).
Since the probability is two-thirds that a particular kernel selected for planting is heterozygous for the gene of interest, the likelihood that any particular pollen parent plant is heterozygous, or that any particular female parent plant is heterozygous, is two-thirds. When a pollen parent plant is self-pollinated and crossed onto a female parent plant that is also self-pollinated (double-pollinated), then the likelihood that both parents are heterozygous is 2/3 x 2/3 - 4/9 or 0.444 (see Coe, MNL 51:61, 1977, for similar considerations). If one pollen parent plant is selfed and crossed onto three different female plants, the probability that the pollen parent and at least one of the female parents crossed by it will be heterozygous is 0.642 (the probability of failing to achieve this situation is 0.358). If three pollen parent plants are selfed and each is crossed onto three different female plants, then the probability of failing to achieve an unambiguous test where both the pollen and female parent are heterozygous is 0.046 (=0.358 x 0.358 x 0.358) and therefore the likelihood of successfully conducting the allelism test is 95.4%. This requires a total of 12 plants, three to be used as pollen parents and nine to be used as female parents for double-pollination. If four different pollen parent plants are used and each is crossed onto two female parents that are double-pollinated, then the probability of a successful test is 97.3%, and this system also requires a total of 12 plants. If six different pollen parent plants are used and each is crossed onto one female parent plant that is double-pollinated, then the probability of a successful test is 97.1%, and this system also requires a total of 12 plants. The choice of which of these schemes to employ (assuming the desire for at least a 95% probability of successfully conducting the test) will depend on the number of plants of both the pollen and the female parent families that are available for pollination.
William F. Sheridan and Janice K. Clark
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}