The two major zein protein size classes of 19,000 and 21,000 daltons are synthesized and stored in approximately equal amounts in maize endosperm. Although the synthesis of proteins of each class appears to be closely coordinated, several important differences have been found in the organization and expression of the two gene classes. The gene clusters for the 19,000 and 21,000 dalton zeins are located on different chromosomes, the cDNAs specific for each class do not cross hybridize, and regulatory loci such as opaque-2 and floury-2 affect the two classes differently.
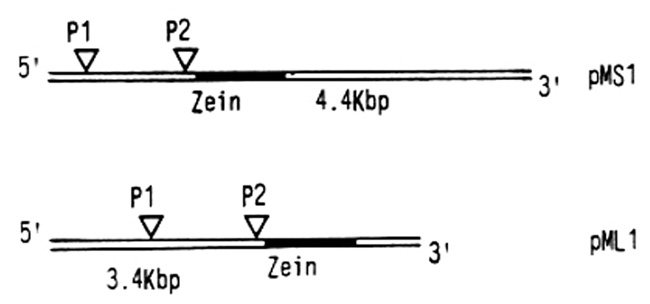
Since the isolation of maize genomic clones containing intact zein genes and extensive flanking sequences, it has been feasible to analyze some of the differences between the genes coding for the two zein protein classes and their mechanisms of expression. As a representative of the 19,000 dalton protein class, we have analyzed the maize clone pMS1 (with a 4.4 Kb maize insert) and as a representative of the 21,000 dalton protein class the clone pML1 (with a 3.4 Kb maize insert). Both classes contain one zein gene free of intervening sequences (as with all zein genes studied to date), and at least 1.5 Kb to the 5' side of the protein coding sequences. The 5' regions have been sequenced and RNA isolated from maize endosperm has been used to accurately map the zein mRNA start positions. These experiments have been combined with in vitro transcription data to demonstrate that both genes are transcribed from two widely separated promoter regions, as indicated in the diagram below.
The two promoters, termed P1 and P2, are separated by about 1,000 bases with no protein coding function. Furthermore, although the positions of P1 and P2 relative to the zein protein coding sequence are very similar in pML1 and pMS1, the sequences of the entire region are different. The sizes of the mRNAs that are transcribed from each promoter correspond in size to the zein mRNAs isolated from endosperm. It is thought that the larger zein mRNAs (over 2,000 bases long) are transcribed from promoters that lie still further 5' of P1.
In addition to the analysis of the transcription of pMS1 and pML1 zein genes in maize endosperm, we have studied the expression of these genes in vitro and in heterologous in vivo systems. The activity of the P1 and P2 promoters for these two genes is summarized in the table below.
The two clones show quite different activities in the different systems. pMS1 was inactive in both in vitro systems but was able to synthesize zein protein, detected by immunofluorescence, in Acetabularia. Conversely, pML1 was active in the in vitro systems but only gave inconclusive results in Acetabularia. Although both clones were actively transcribed in yeast, pMS1 used exclusively whereas pML1 only used P2. The RNA starts used in yeast were identical, however, to the nucleotide, with those used in endosperm.
The ability to transcribe and, as in the case of Acetabularia, to translate the zein genomic clones greatly increases our ability to study the control of expression of these genes. The clear-cut differences in the expression of the two genomic clones, both in terms of absolute activity and with respect to the activity of the two promoters, throw some light on the differences observed in the expression of the two zein protein classes in endosperm. Such differences may, in fact, lie behind differences in action of mutated regulatory loci. With the heterologous systems that we now have in use, we hope to study more closely the signal structures associated with zein gene expression. More definitive studies must, however, await the development of a homologous transcription system.
P. Langridge, J. Brown and G. Feix
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}