From a population of Peruvian maize, cultivated in greenhouse conditions, one plant was pollinated by the French early line F7. In the offspring of this hybrid, a plant (P2) was recognized as susceptible to methomyl by leaf test bioassay. By backcrossing this material by F7 or F215 inbred (as male recurrent parent), some lines were obtained that were remarkable in several aspects:
1. After three backcrosses by F7, most of the progenies appear male-sterile and susceptible to HmT (Helminthosporium maydis race T) toxin and methomyl. These traits are characteristic of Texas cytoplasmic male-sterility. However, the male fertile plants obtained among the segregants did not restore the male sterility of F7-T line.
2. Part of this material, crossed at least 2 times by F215 (cms-T maintainer), gave different results: most of the plants were male-fertile and segregated for resistance/susceptibility to HmT and methomyl.
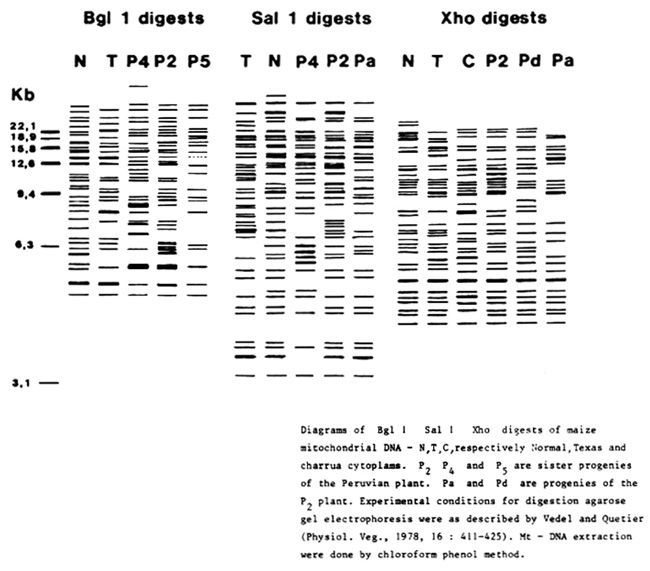
3. A mtDNA analysis of P2, P4 and P5 hybrid progenies (Peruvian X F7) and several offspring from P2 has been started, according to the methodology described by A. Berville and M. Paillard (Les Colloques de VI.N.R.A., 11:125-136, 1982). BglI, SalI and Xho restriction enzymes were used as shown in the figure. Obviously, P4 exhibits a pattern, whatever the enzyme used, never described for maize mtDNA: the two large bands at 9 and 5.5 Kb are typical. P5 looks like T after digestion by SalI (not shown) but appears different after digestion by Xho (see figure); furthermore, P5 is resistant to both methomyl and HmT toxin. In contrast, P2, which is susceptible to both compounds, exhibits a pattern that is quite different with the 3 restriction enzymes. Moreover, P2 progenies, Pa and Pd, for example, are different from the P2 origin (with Xho and SalI), which in several aspects look like N patterns (BglI and SalI). However, in other progenies showing the same phenotypes, P2 pattern does not appear to be modified.
It is too early to explain why in this population the mtDNA is so unstable. The cross by the French line may have initiated rearrangements or partial amplifications within the mitochondrial genome. The mtDNA analysis of our material from backcrossing by F7 or F215 could permit us to establish whether the male parent is involved in such mtDNA variability. Nevertheless, it seems evident that in this maize population there could coexist several patterns of mtDNA, similar to or different from the T, C, N (see figure) or S (not shown here) types previously described.
A. Mezzarobba, J-C. Nicolas, A. Berville and A. Cornu
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}