This report concerns variants discerned among the set of polypeptides peripheral or intrinsic to the chloroplast thylakoid membrane of the genus Zea. The phenotypes are detected by means of a system of gradient-density SDS gel electrophoresis, based on modified Laemmli buffers and with the addition of urea to the resolving gel. The density range of the resolving gel, the pH resolving buffer, and the amount of SDS (added to the cathodic buffer) are varied in order to optimize the resolution of the particular migrational variant. Proteins are visualized by staining with Coomassie Brilliant Blue R-250. Apparent molecular weight references are relative to the migratory positions of the six proteins composing the Low Molecular Weight Standards, offered by Bio-Rad Laboratories, and hold true only within the running conditions used to visualize the particular protein of interest. A precise protocol is available from the author (c/o E. H. Coe, 210 Curtis Hall, UMC, Columbia, MO 65211).
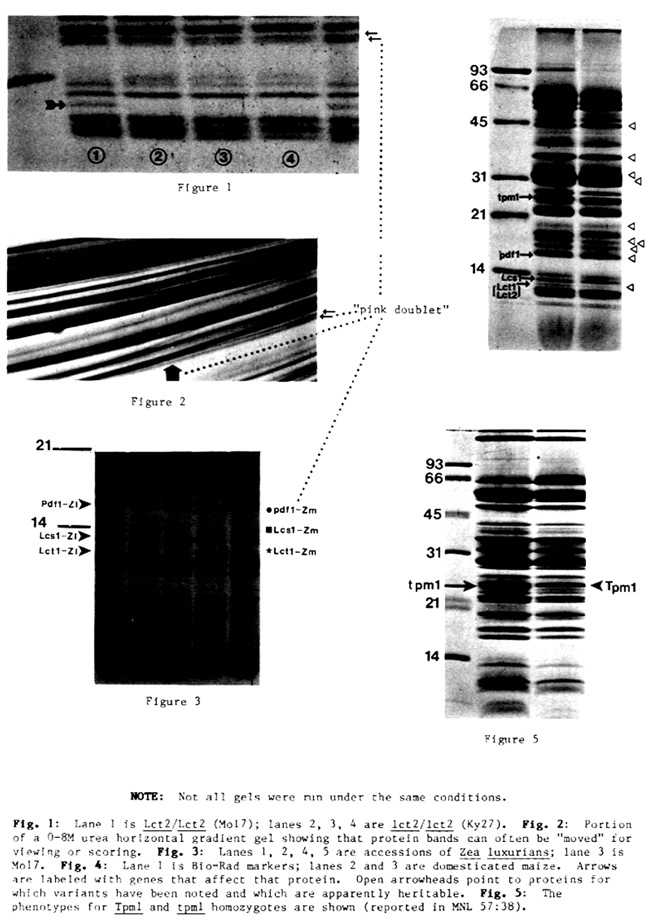
For the sake of brevity, detailed accession information and gel photos of variants are omitted from this report. It can be obtained from the dissertation of Stephen Anthony Modena (upon its acceptance) or by direct communication. A gel profile of "wild type" maize is included and labeled for the general orientation of those working with chloroplast proteins.
I have adopted the practice of naming these genes in a trivial manner. I do not know the function of the proteins involved. Their exact relative migratory location on a gel will depend on the interaction of gel running conditions. The electrophoretic conditions used in some laboratories will not even resolve these bands, let alone the variants. Positive identification and further characterization of specific variants will depend on pedigreed stocks. Small quantities of seed, intended for increase first, are available from the author upon request.
I can resolve a cluster of at least seven bands, the slowest migrating of which approximately aligns with the 14.4 Kd. standard. I count from this slowest band.
Lcs1: For the second slowest band of the cluster I have detected three migrational phenotypes: wild-type domesticated maize has the fastest form; Z. luxurians, the slowest; and Z. diploperennis is heterogeneous for an intermediate migrational form and a maize mimic migrational form. Our one accession of Z. perennis is maize-mimic. A limited survey of Corn Belt and exotic germplasm of Meso-America uncovered no other alleles in maize. A cursory survey of Balsas, Chalco, Huehuetenango, and Nobogame indicate that they possess the maize phenotype.
For Z. diploperennis-maize reciprocal crosses, the individuals conform to the phenotype of the Z. diploperennis parent involved. In the progeny of the lineage (((Z. d. x Z. m.) x L. m.) x Z. m.), in which the Z. diploperennis parent showed the Z. d. slower form as did the first backcross ear parent, the parental phenotypes are segregating. A symbol or allele is not yet assigned because statistically insufficient progeny have been scored.
Data are presented for several crosses involving Z. luxurians:
One gene or two? Or is there a biasing effect here related to the fact that the interspecific F1 plants characteristically are two-thirds pollen aborted? The quality of the Lct1 data (below) is a contrast!
In the belief that at least one nuclear factor is involved, the symbol Lcs1 (last cluster, second down) is assigned and there are two alleles: Lcs1-Zm and Lcs1-Zl. I can not resolve the genotype Lcs1-Zm/Lcs1-Zl into two bands, but there is band broadening uncharacteristic of either of the homozygotes. I favor the idea that this is structural variation.
Lct1 and Lct2: For the third slowest band there are two genes affecting the phenotype: structural variation and a presence-absence conditioning gene.
All Z. luxurians accessions examined possess a faster migrating form of this protein band; all domesticated maize (with two exceptions) and the teosintes (Balsas, Chalco, Huehuetenango, Nobogame, Z. diploperennis, and Z. perennis) examined have the wild type maize form. The single cross, maize x Z. l., has two bands of approximately half intensity; one aligns with maize, while the other aligns with Z. l.
Drawing upon exactly the same progeny as for scoring the Lcs1 gene, segregation data are presented in the following table.
The gene symbol Lct1 (last cluster, third down) is assigned and there are two allelic forms: Lct1-Zm and Lct1-Zl.
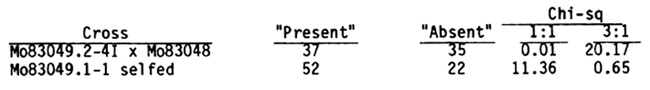
The second locus affecting the expression of this band was recovered from the inbred line Ky27 (See accompanying article for a fuller story.) This was readily detected because the band of interest is apparently absent. Though various running conditions have been tried, I have not been able to detect a band at a position away from maize wild type. Nor is there any indication of an adjacent or nearby band intensifying, indicating migrational overlap--not in the F1 or the F2. Because of the apparent dominance-recessive nature of the segregation, this is unlikely to be another version of Lct1. The test cross and F2 data are presented:
The gene symbol lct2 is assigned and there are two alleles: Lct2 and lct2-Ky27. The inbred Ky122 has a gel phenotype like Ky27, but an allelism test has not been done.
The "absence" of this protein in lct2 plants raises at least two interesting possibilities: 1) this is a "dispensable" component of the thylakoid membrane or 2) there is another protein in the membrane (at a different migrational position on the gel) which performs a homologous function, affording the Lct1 protein to be absent.
Others wishing to work with these genes are welcome! Better techniques and fresh approaches may resolve some of the above ambiguities. Meanwhile I am converging these genes to some standard maize backgrounds.
Stephen A. Modena
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}