Many of the hcf mutations isolated by C. D. Miles and coworkers have been tentatively assigned to chromosome arms by B-A translocation analysis. Some preliminary linkage data to mapped markers are given below. The hcf stocks employed were obtained from Miles and used directly, or were subjected to one or more successive crosses with the inbred Mo17. Stocks with multiple markers were obtained from the MGC Stock Center.
hcf*-9 (allelic to hcf*-3): F2 progenies that segregated hcf, sr and zb4 in a given progeny were obtained by crossing +/hcf*-9 individuals with pollen from a + + P-WW/sr zb4 P-WW plant. Data for one progeny are given in Table 1, and show
hcf*-9 is linked to zb4.
All traits did not segregate at 25% in progenies where they were observed, preventing estimation of recombination distances. Segregation for the progeny described in Table 1 was as follows: zb4, 19.9%; sr, 24.9%; hcf, 29.8%. Of all sr individuals referred to in Table 1, 24.4% were also hcf, and of the zb4 individuals, 14% were hcf. The low number of members of two recombinant classes with hcf prevent establishment of a gene order.
Comments on expressions: 1. Night temperatures of 70 F did not give good expression of zb4. After emergence of the first leaf, placing seedling flats overnight at 50 F produced a fine but distinct yellow green horizontal band on leaves after further growth. Similar banding could not be produced for progenies segregating hcf*-9 only. The number of fine bands on each leaf usually corresponded to the number of nights material was kept at 50 F. 2. The hcf trait was scored by counting lethals, many of which had been confirmed to be hcf. The high number of "hcf's" may thus be due to lethality for other reasons, e.g. poor vigor of sr individuals with large amounts of albino tissue. In the future, hcf individuals will be scored by fluorescence induction.
hcf*-6: F2 progenies segregating hcf*-6, sr and zb4 are being analyzed.
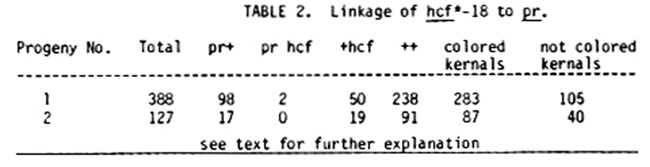
hcf*-18, hcf*-43: Summer 1982 crosses indicated that hcf*-18 and hcf*-43 are allelic. The low number of pr hcf individuals that segregate in the F2 progenies a cross between +/hcf*-18 and +++/bm pr ys is evidence that hcf*-18 is linked to pr (see Table 2).
Two problems with this test prevent establishment of gene order of hcf*-18 relative to pr, ys, and bm: (1) the hcf phenotype only occurred in 13-15% of the individuals for several F2 progenies examined (as assessed both by lethality of 2-3 week-old seedlings and by induction of fluorescence). Not all progenies segregating hcf*-18 or hcf*-43 have this property. Future linkage tests will be repeated with several +/hcf*-18 or hcf*-43 individuals from progenies which segregate 25% hcf seedlings. (2) ys could not be scored unambiguously in unfertilized seedling flats, and bm expression required seedlings where at least 4 leaves had fully expanded. The hcf seedlings generally died sometime before the third leaf had expanded. The F3 generation is being examined for segregation of hcf in ears that are ys/ys, or bm/bm or Pr/pr or combinations thereof.
Other hcf markers being tested for linkage, but where F2 progenies have not yet been generated, are (the numbers in parentheses are tentative chromosome arm assignments provided by C. D. Miles): hcf*-2 (1L), hcf*-4 (1L), hcf*-12 (1L), hcf*-13 (1L), hcf*-41 (1L), hcf*-44 (1L), hcf*-50 (1L), hcf*-21 (5L) and hcf*-42 (9L). All of these hcf mutations have either very distinct effects on photosynthesis or have been shown to be nonallelic.
Mary Polacco
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}