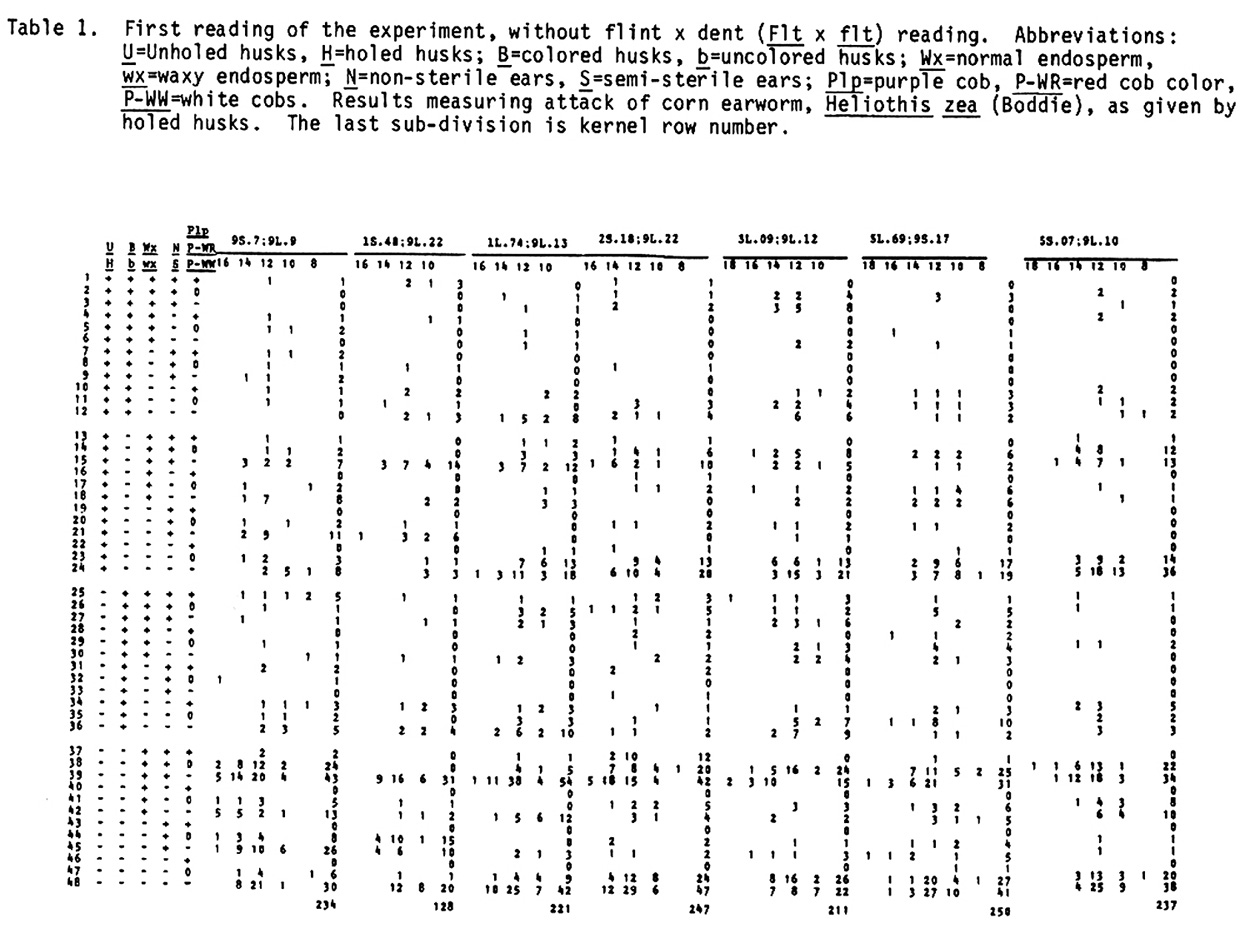
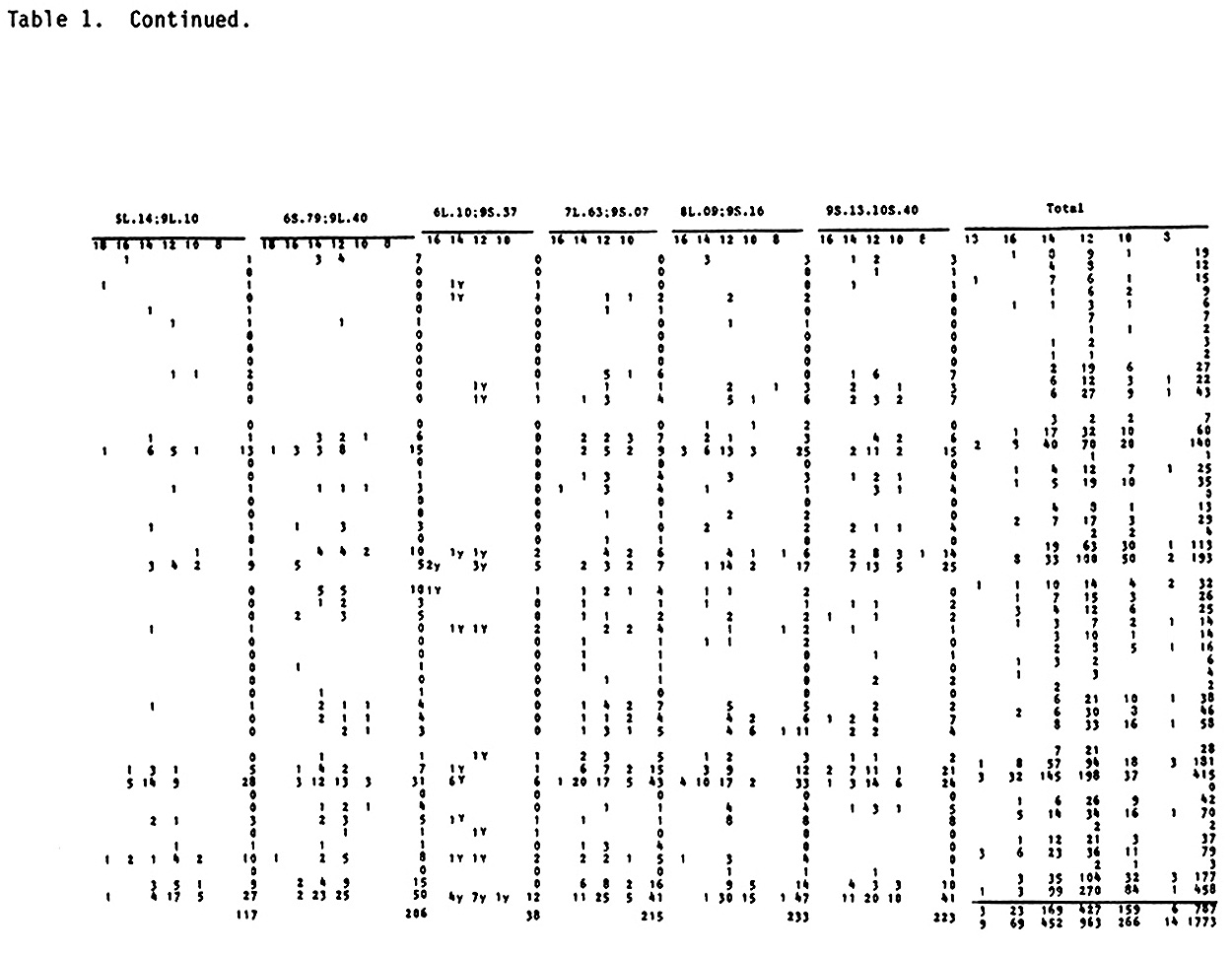
The authors (MNL 56:30-32 and 57:20-24) reported results on horizontal resistance with the standard 9 wx translocations in IAC Maya lte1 crossed to Zapalote Chico (ZC) and from bibliographic research. Now we report results with the same markers crossed to Cateto Palha Roxa (Purple Husk Cateto). This is a backyard collection picked in the Paraiba Valley between Sao Paulo and Rio near the city of Pindamonhangaba. It is a pure Cateto maintained by a conservative farmer family for generations and is reported to be resistant to weevils. The cross was "back-crossed" to IAC Maya wx y. The field plots consisted of one row 10m long planted in hills at 0.4m within the row. The hills were planted with three seeds each, alternating hills with normal and waxy endosperm of the same ear within the row. Six reps were used. Every fifth row was planted with IAC Maya wx y as pollinator, and the experiment was detasseled. The dry ears were harvested indiscriminately, unhusked. They were classified for presence vs. absence of exit
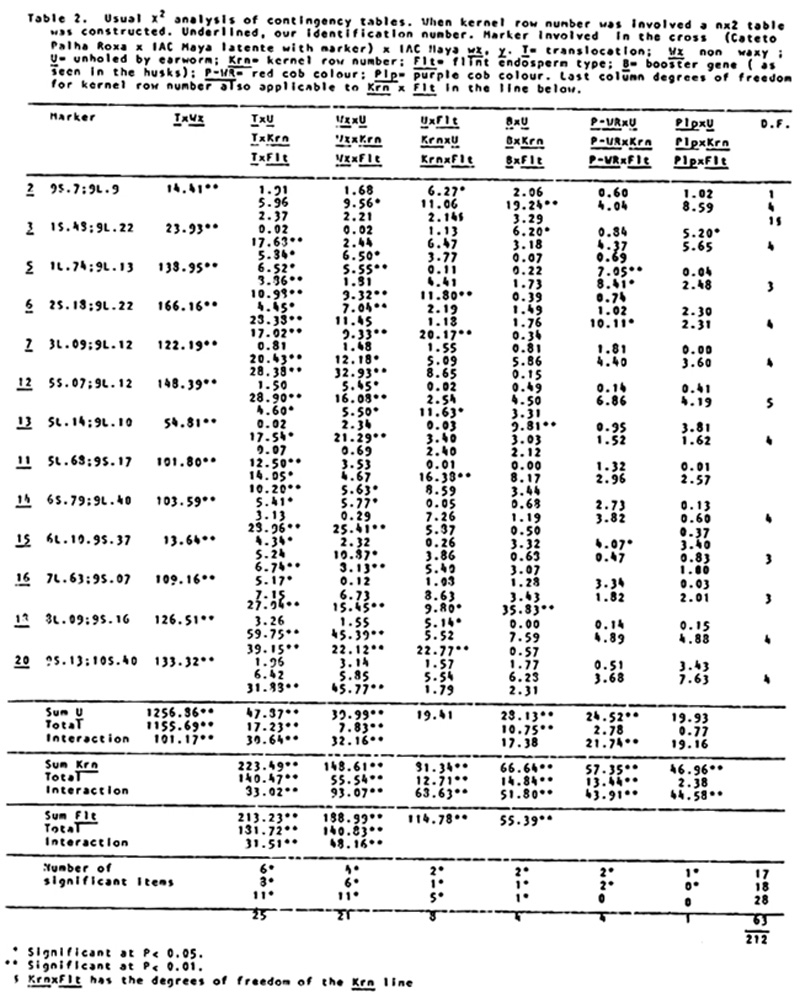
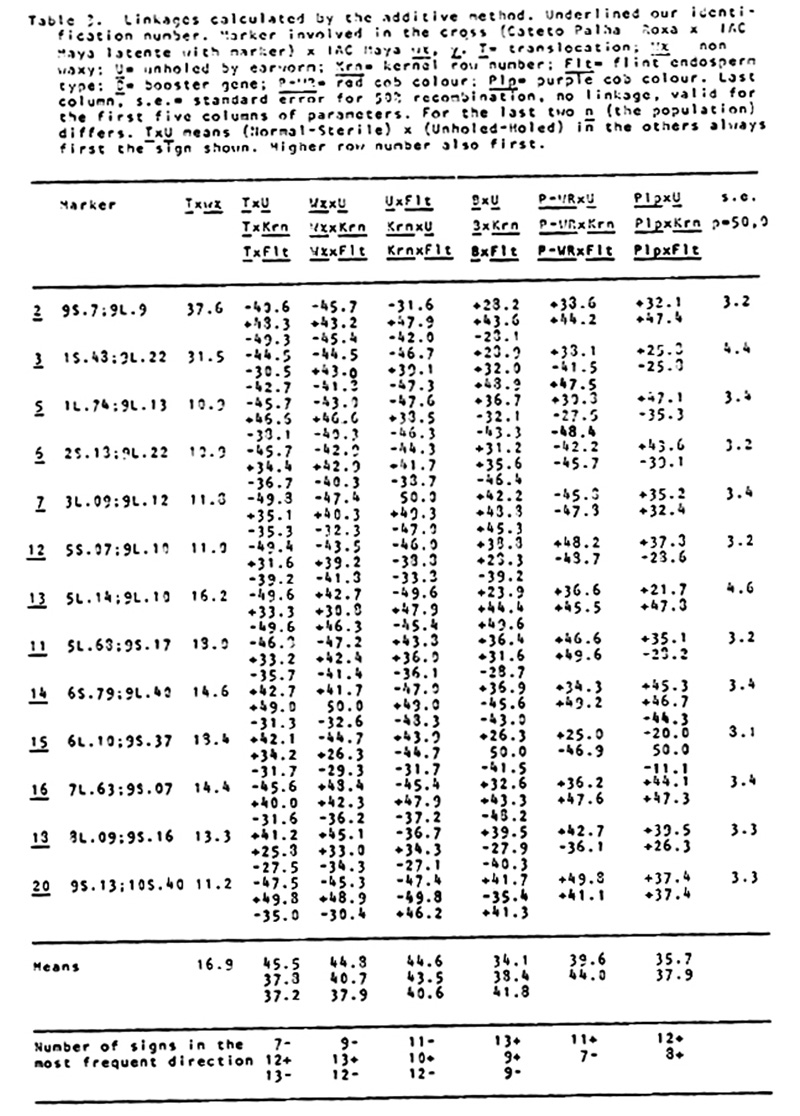
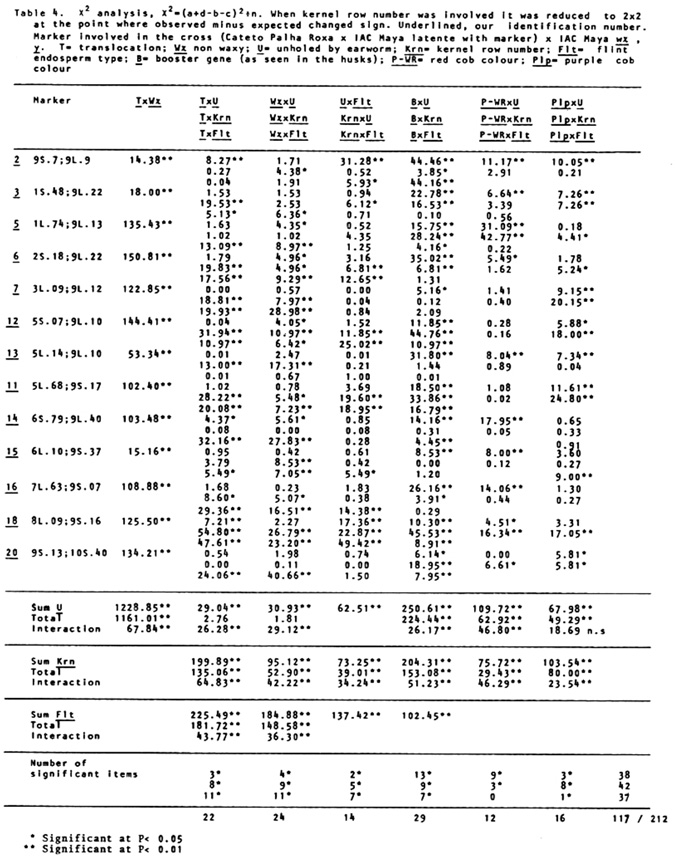
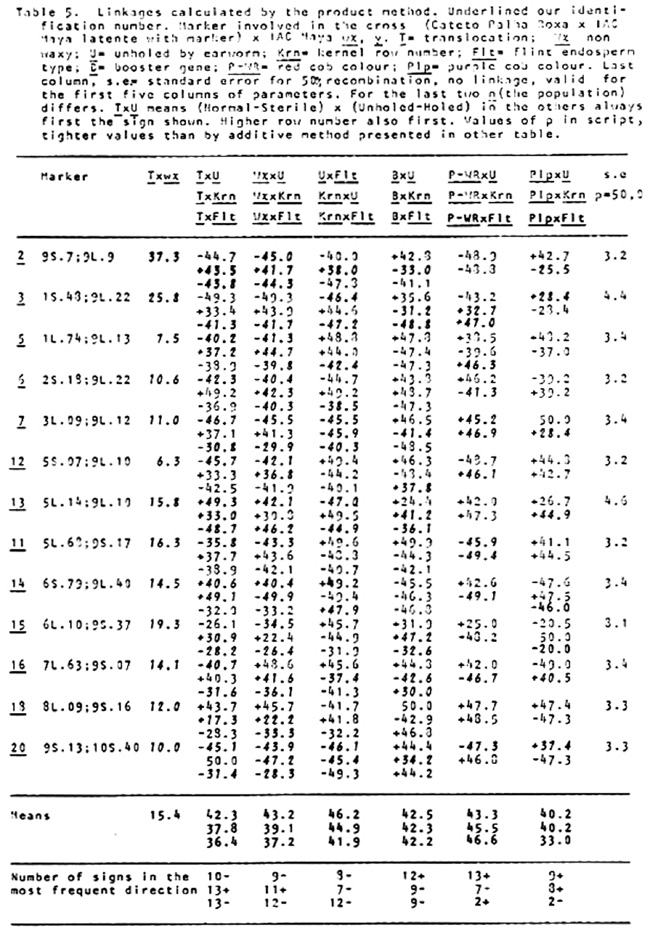
holes of corn earworm, and for degree of damage in the cob by an adaptation of N. W. Widstrom's scale (J. Econ. Entomol. 60(3):791-794, 1967), lumping the 0 and 1 class in one and from two forward in the other, or without visible damage and damaged in the cob. Besides normal and semi-sterile ears, and Wx and wx, since the Cateto is P-WR and B-W and has a high frequency of Plp (Purple restricted to the cob), these factors were also classified, and the kernel row number was counted. A preliminary analysis was done of the two methods of measuring attack of earworm, unholed and holed husks versus undamaged and with visible damage in the cob. The second method gave lower X2 values and was henceforward dropped from further analysis. Data from the first reading are given in Table 1. A X2 analysis in the usual way is in Table 2. In Table 3 are the values of p calculated by additive method, always in the direction of linkage. It is evident in the linkages and X2 values between wx and translocation that there are quite disturbed segregations. Consequently a second X2 analysis was done using the contrast (a+d-b-c)/n, squared (Table 4). The values obtained compared with the first method show that this method is also quite indicative of linkages. So a second calculation of p was done using the product moment method for backcross, equating (l-p)2/p2 = (axd)/(bxc) which resolves into the quadratic (axd-bxc)p2+2bcp-bxc (Table 5). The standard error is the same as by maximum likelihood, which is equal to the additive method in this particular case. In general the product moment method was more efficient, giving tighter values of p when the translocations were involved. All p values between the translocations and wx were tighter. For further analysis it was considered the stronger value between the two methods. With these values we applied the theorem: the real distance in centimorgans between two factors with heritability less than one is the difference between the distances of them converted to centimorgans with a third factor outside their span. As illustrated by cross number 5 (2 and 3 were quite abnormal):
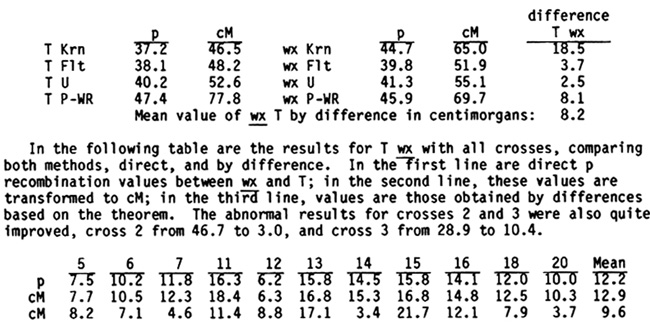
In the following table are the results for T wx with all crosses, comparing both methods, direct, and by difference. In the first line are direct p recombination values between wx and T; in the second line, these values are transformed to cM; in the third line, values are those obtained by differences based on the theorem. The abnormal results for crosses 2 and 3 were also quite improved, cross 2 from 46.7 to 3.0, and cross 3 from 28.9 to 10.4.
The first analyses were done without taking readings for the flint (Flt) character. On the stored material a second reading of the data was done for flint x dent dividing visually in two classes (Table 6). For the appropriate chromosomes a reading was done of Flt with P-WR, B-W and Plp. Since Cateto is Bn and IAC Maya wx y has a high frequency of bn, this factor was also read accordingly. From these last readings only the p values are presented. With the method above illustrated by linkages between translocations and waxy, all combinations of linkages between factors were calculated among all factors. With these values were mounted the tentative linkage groups listed after Table 6, with positions and distances indicated.
We have also taken into consideration the Cytogenetic Working Map in MNL 52:129-145. For linkage of Flt8 in chromosome eight we had unpublished data in crosses of Cateto with 8L.09;9S.16 and 8L.35;9S.31, showing more linkage with the latter in a more distal position of the long arm.
Since for earworm resistance we had used symbols Zer4 and Mer2, we continued to identify them with glucoside earworm resistance symbol (Ger) starting from one.
With the possible exception of chromosome 10, which had only an Flt factor, all the others have factors for Flt, Krn, and Zer, Mer or Ger.
As seen by the results, with the possible exception of chromosome 6, the Krn and Flt factors are always adjacent to one another, with Zer, Mer or Ger outside their span. Also these groups are near chlorophyll and endosperm characters, meaning that the inheritance of horizontal resistance, kernel row number, and finally carbohydrate synthesis, are governed by overlapping super-genes.
L. T. de Miranda, L. E. C. de Miranda, C. J. Rossetto and Eduardo Sawazaki
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}