In 1982 and 1983 Stuber and Goodman (MNL 56:127-132, 1982; USDA-ARS, ARS-5-16:1-29, 1983) presented genotypes for 21 enzyme loci in 406 U.S., Canadian, and European inbred lines of corn. Since 1980, we have also routinely characterized public inbred lines of maize for their genotypes at polymorphic enzyme loci. Assays have been restricted to loci which could be read unambiguously with electrophoretic conditions used in our laboratory (Crop Sci. 23:572-576). All genotypic information included in this report comes from inbred lines obtained from other stations. The inbreds are being maintained and used in population genetics and population improvement research at the Northern Grain Insects Research Laboratory at Brookings. Genotypes of these inbreds are given to complement the earlier lists provided by Stuber and Goodman.
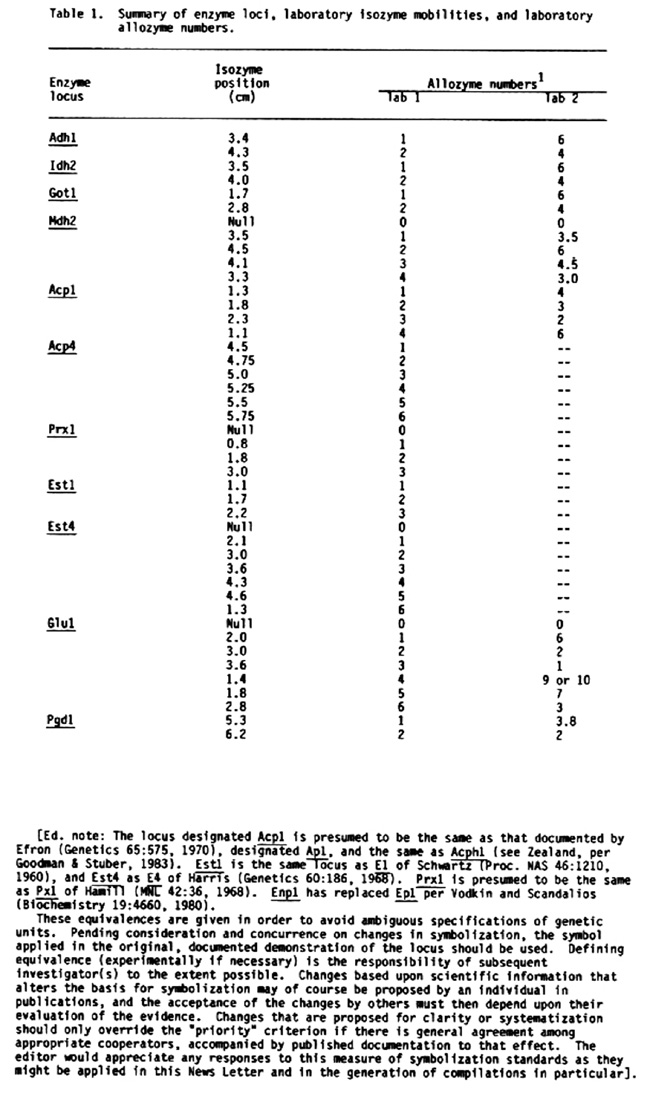
Electrophoretic methods have been reported for all loci assayed (Cardy et al., Inst. of Stat. Mimeo. Series No. 1317, N.C. State Univ., 1980; Kahler, Crop Sci. 23:572-576, 1983). Enzyme loci, laboratory isozyme mobilities from the gel origin and laboratory allozyme numbers are given for convenience in Table 1.
Corresponding laboratory allozyme numbers of Stuber's laboratory are also given in Table 1 for all cases where the correspondence has been determined.
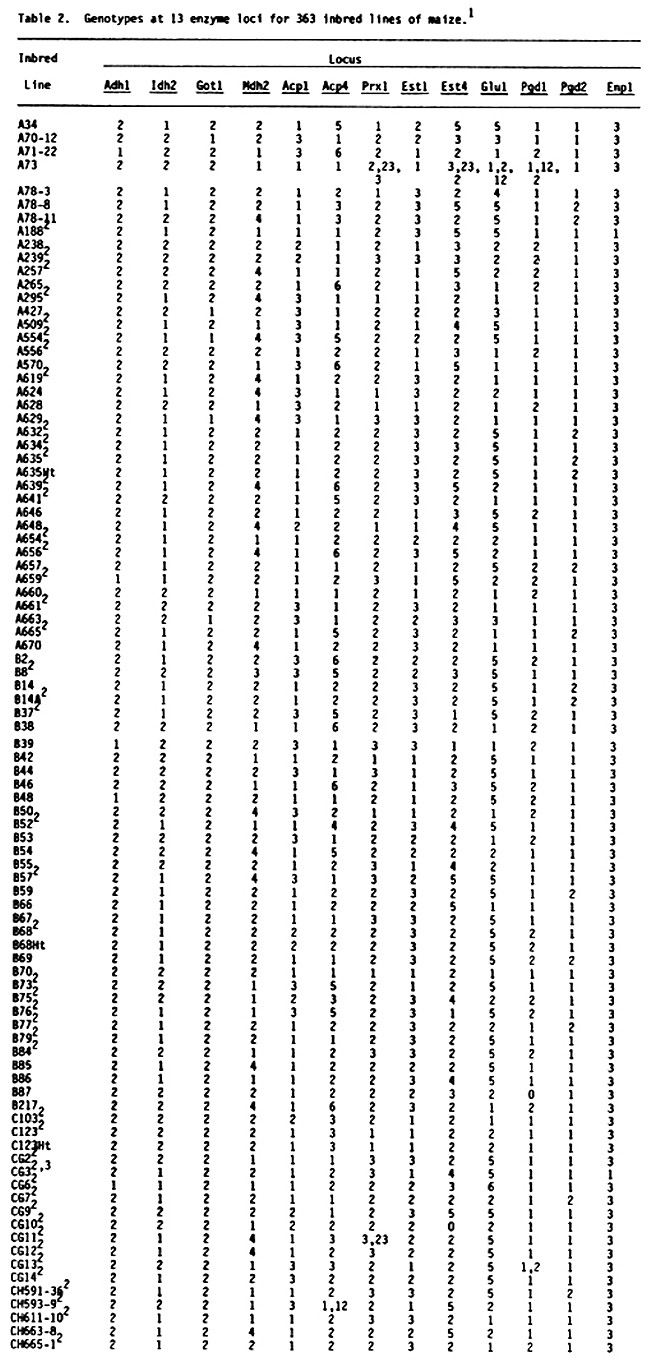
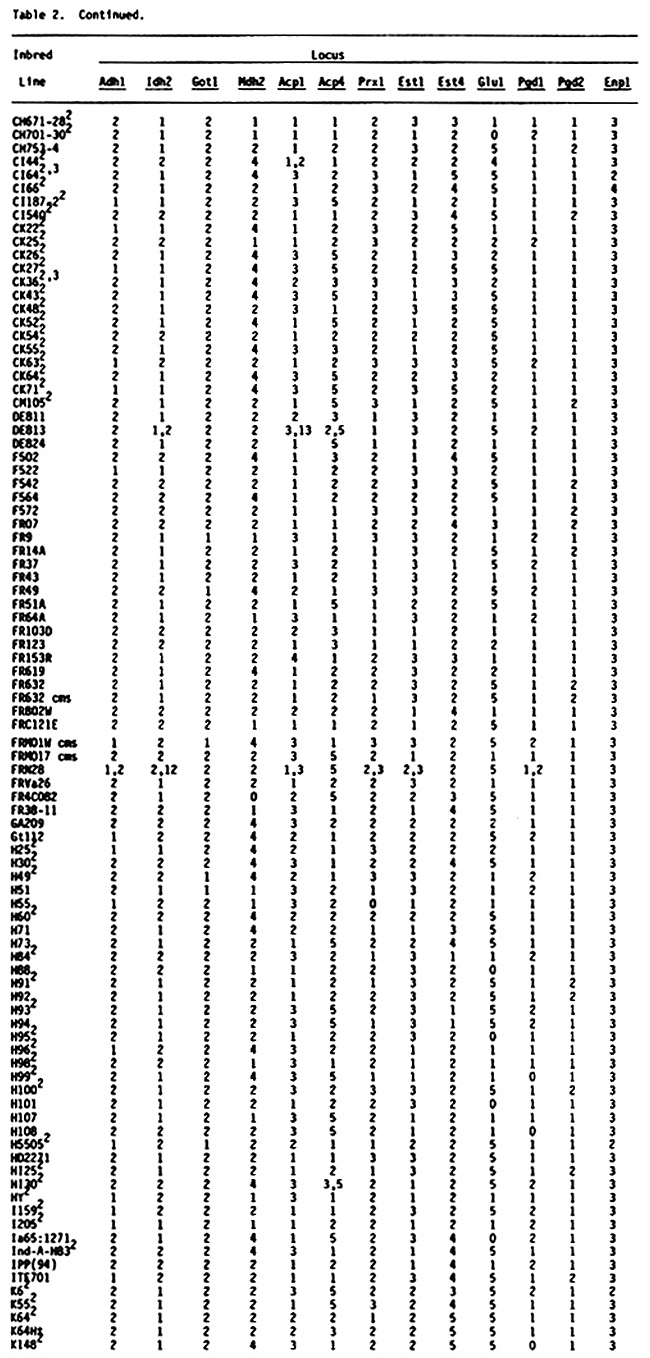
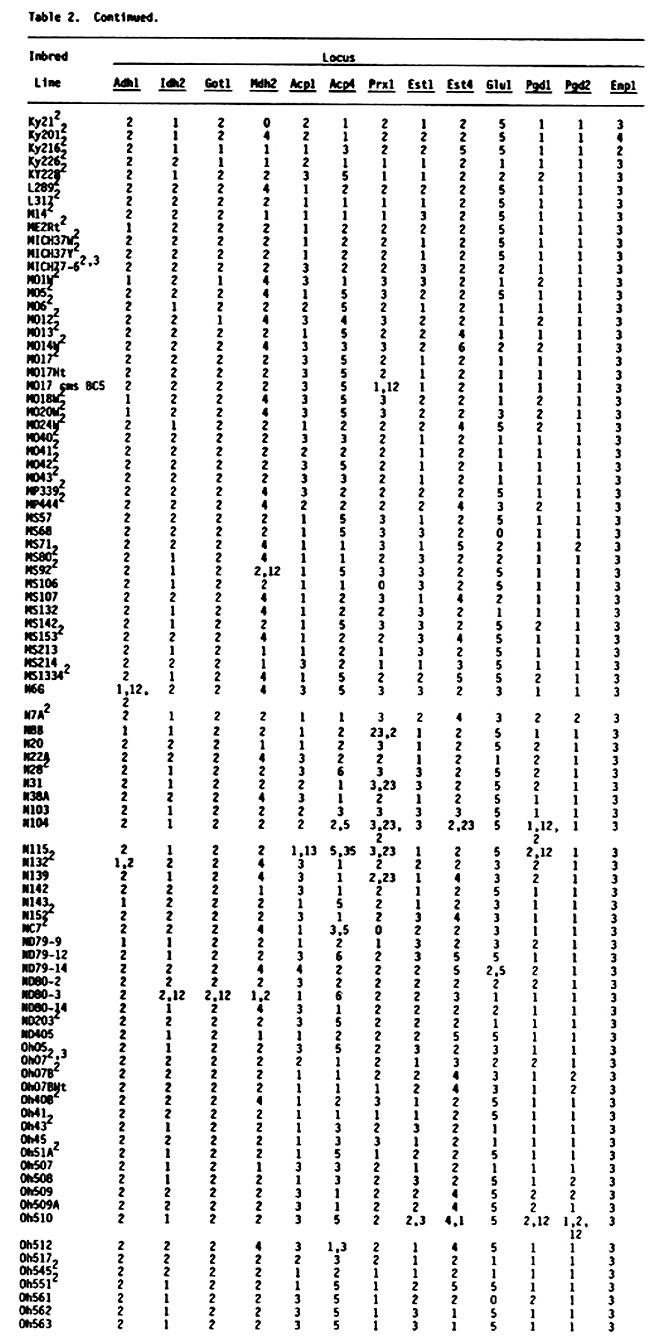
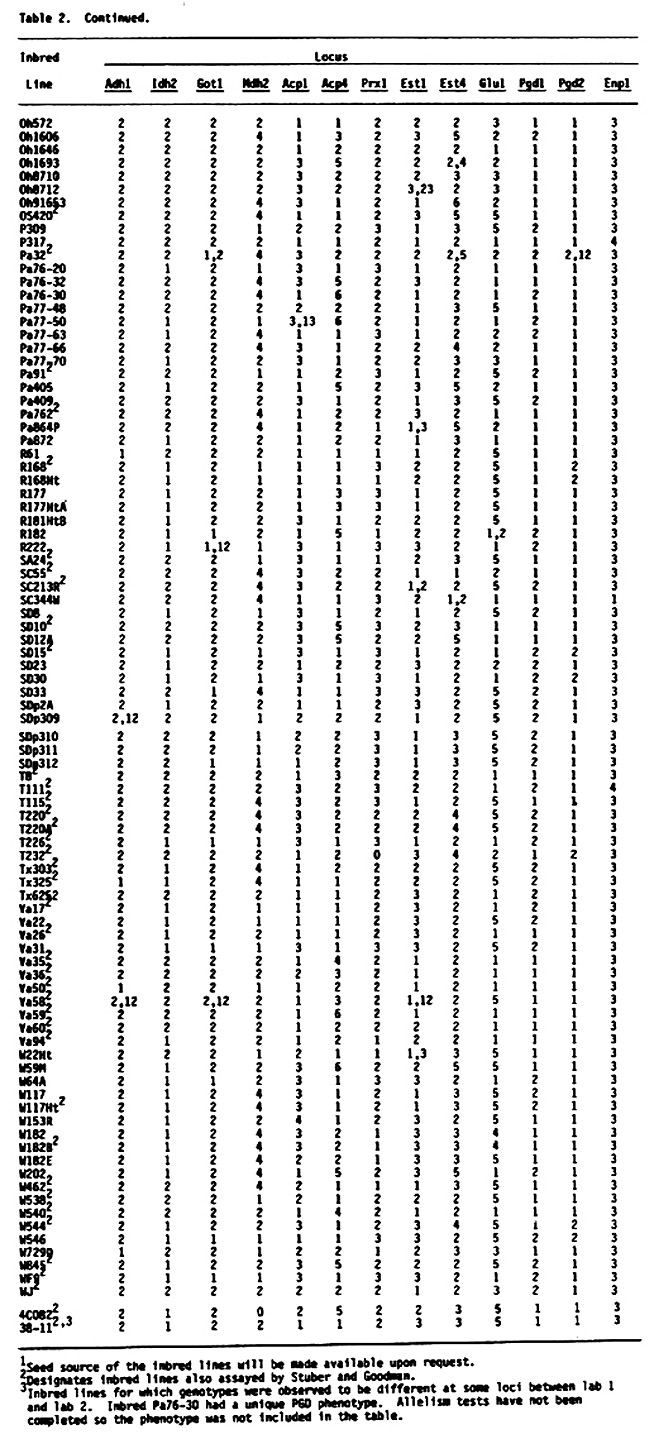
Table 2 (see preceding pages) presents genotypes at 13 enzyme loci (Adh1, Idh2, Got1, Mdh2, Acp1, Acp4, Prx1, Est1, Est4, Glu1, Pgd1, Pgd2, Enp1) for 363 U.S. inbred lines of maize. One hundred seventy five of these lines are common to the earlier lists, although the source of common lines may be different. These 175 lines are included because genotypes at loci Acp4, Prx1, Est1 and Est4 were not included in earlier lists. Genotypes are usually designated with a single number in the table because most lines are homozygous and homogeneous. For example, genotype 2 at locus Adh1 symbolizes the Adh1-2/Adh1-2 homozygote. Heterozygotes are designated with two numbers representing the allelic state of the hybrid. Thus genotype 12 at locus Adh1 symbolizes the Adh1-1/Adh1-2 heterozygote. When a line was heterogeneous all observed genotypes were presented with the most frequent genotype first, followed by the next most frequent genotype and so on. A minimum of two, and as many as 40, individuals were assayed per inbred line. Because of the relatively small sample sizes for some of the inbreds, rare variants probably were not detected. Therefore, these data should be considered preliminary. It is possible that some of the "uniform" inbreds are polymorphic due to contamination from seed mixing or outcrossing. In addition, genotypes for some inbreds and loci differ from earlier reported genotypes. In most cases this reflects different seed sources. Seed sources for inbred lines included in Table 2 will be made available upon request.
Among the 594 inbred lines that have been characterized for genotypes at enzyme loci, it is now possible, by inference, to determine expected single-locus and multilocus genotypes of 176,121 possible single-cross hybrids.
Table 2. Part 1. Part 2. Part 3. Part 4.
A. L. Kahler, J. Matthees and R. Telkamp
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}