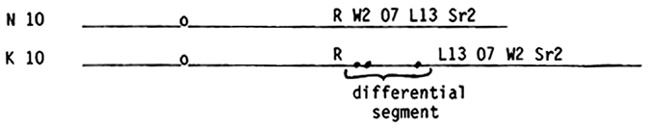
In a series of reports in the Maize News Letter, we demonstrated by cytogenetic methodology that the distal portion of the long arm of abnormal chromosome 10, which is found in certain races of maize grown in Arizona, New Mexico, Mexico and the Central American countries, differs drastically from normal 10 in its architecture, amount of chromatin, and linear order of genes. Our conclusions are diagrammatically summarized below:
In both kinds of chromosome 10 the W2 07 L13 segment is situated between the R and Sr2 loci, but in abnormal 10 this segment has undergone excision and transposition distally in an inverted order with respect to the flanking R and Sr2 genes. A piece of chromatin of unknown origin possessing three small knobs, designated as the differential segment, lies interposed between the R and L13 loci. A second piece of chromatin, again of unknown origin, composed of a large heterochromatic knob with a short euchromatic tail, terminates the long arm. We suggested that a minimum of three breaks occurred in the origin of abnormal 10 from a normal 10. One break was between R and W2, a second between L13 and Sr2, and the third between Sr2 and the telomere. One end of the differential segment, carrying no marker genes, was inserted next to the R locus with the other end joined to the L13 end of the W2 07 L13 segment. The order of loci in this segment is inverted relative to their orientation to R and Sr2. Finally, a second piece of chromatin, origin unknown, comprised of a large heterochromatic knob and a short euchromatic tail, was inserted subterminally just distal to Sr2, giving the structure of abnormal 10 shown above. The fate of the normal telomere is uncertain; it may be present as the telomere of the long arm of K10 or it may have been eliminated.
Before accepting this complex, even bizarre, scenario we should critically examine the various links in the chain of argument to see if some are of questionable validity. For example, the statement that the L13 07 W2 block of genes is inverted in the transposed location in abnormal 10 assumes that the order in this segment in normal 10 is R W2 07 L13 Sr2. This is based on the order determined from the collated linkage data given on page 191B of the 1983 Maize News Letter, where the order and map positions of loci distal to R are as follows:
As is well known, backcross data provide more precise recombination values than do F2 data in coupling, and much greater precision than do F2 data in repulsion. Homozygotes for either w2 or l13 are lethal, so backcross data cannot be obtained and F2 data must be resorted to in locating these two genes. Furthermore, homozygous l13 or w2 seedlings cannot be classified for the sr2 phenotype, and w2 is epistatic to l13, thereby increasing the difficulty of determining map positions from F2 data. An additional uncertainty is the variation in recombination values for the same interval in different strains of maize. Since the location of w2 was based on F2 data, and since our argument was so dependent upon ascertaining the correct linear order in the distal portion of the long arm of both normal and abnormal chromosome 10, we decided to undertake the arduous task of genotyping every plant in a population segregating for R, w2 and sr2. This is a laborious process since every tested plant gives the genotype of only one gamete but the data are equivalent to backcross data. Mascia (1978 MNL) employed this procedure in accurately placing the l13 locus four map units proximal to sr2. The following cross was made in order to determine the crossover distance between R and W2 (region 1) and between W2 and Sr2 (region 2) in the heterozygous male parent:
The above mating is a testcross for the R and Sr2 loci, while determination of which W2 allele is present in a gamete comes by pollinating all plants with a strain that would reveal the presence or absence of the w2 allele. This permitted identification of the eight genotypic classes listed below along with their frequencies. The chromosome contributed by the pollen parent is given in the denominator.
These data clearly demonstrate that w2 lies between R and Sr2, and give an accurate determination of the w2-sr2 interval. Our data, together with those obtained by Mascia for the R l13 sr2 region, demonstrate that the linear order for 10L given in the 1983 MNL is correct and any misgivings we held about the map positions of these loci were laid to rest. The reverse order of L13 O7 W2 in the abnormal 10 chromosome was unequivocally established by our series of simple terminal deficiencies, since in two independent cases, DfK10(F) and DfK10(I), the W2 locus was deleted but the L13 locus was still present. The data confirm our conclusion that the W2 O7 L13 segment of N10 was both transposed and inverted in the origination of abnormal 10.
M. M. Rhoades and E. Dempsey
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
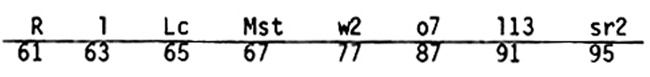{kind=link}
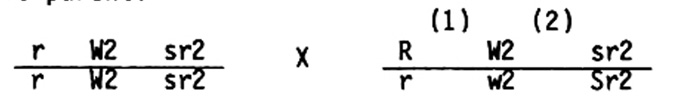{kind=link}
{kind=link}