We reported in last year's Newsletter that the protein products of the Prot locus, PROT' and PROT, account for the major portion of the globulin fraction in maize embryos. Analysis of Prot protein synthesis in vivo and in vitro indicates PROT is derived from PROT', and PROT' is derived from a relatively short-lived precursor, which we have termed prePROT'.
Prot protein synthesis in developing embryos was examined by pulse-chase radiolabeling methods. Intact immature embryos (23-28 days post-pollination) were excised from surface-sterilized kernels, collected under aseptic conditions, and placed on agar culture medium in petri dishes. The basal medium was that of Miller (in Moderne Meth. Pflanzenanal. v.6, Springer-Verlag, 1963), to which abscisic acid was added to 10-5 M; the inclusion of abscisic acid in the culture medium was necessary in order to prevent germination. Embryos which had been conditioned to the medium for two days were pulse-labeled with radioactive amino acids by transferring the embryos to a small petri dish to which either 3H-leucine or 3H or 14C-labeled amino acid mixture was added in the liquid culture medium. Generally, 80-100 microcuries of label in 1 ml of solution were used per 15-18 embryos. The sealed dish containing the embryos was gently agitated during the pulse-labeling period (15-60 minutes). Pulsed embryos were transferred to culture plates and incubated (chased) for 1-72 hours. All incubations were performed at 24 C. The chase period was terminated by either freezing the embryos in liquid nitrogen or dry ice, or by macerating the embryos in a SDS sample buffer and boiling for two to five minutes. Frozen embryos were generally used for immunoprecipitation analysis. Individual embryo extracts were examined by SDS-polyacrylamide gel electrophoresis and fluorography. In vitro translation of immature embryo RNA was performed using both the rabbit reticulocyte lysate and the wheat germ extract systems (BRL).
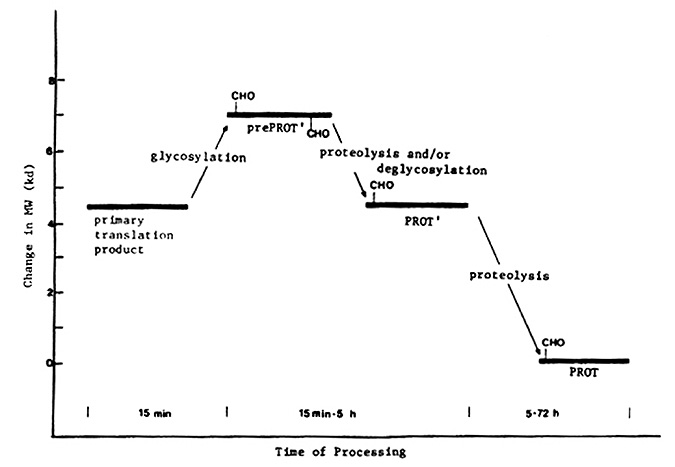
The results obtained from the pulse-chase and the in vitro translation experiments are presented schematically in the accompanying figure. The primary translation product, as determined by in vitro translation of embryo RNA, is approximately 2.5 kilodaltons smaller than prePROT', the initial Prot-specific polypeptide detectable in in vivo pulse-chase experiments. This indicates that the primary translation product is modified co- and/or post-translationally in vivo to yield prePR0T'. Although the nature of this modification is not known, glycosylation is suggested as a probable alternative, as preliminary evidence indicates the Prot proteins are glycoproteins, and other seed globulins are glycosylated in vivo. The processing of prePROT' to form PR0T' may involve deglycosylation as well as proteolysis; this suggestion is based on comparison of prePROT' and PROT' with respect to isoelectric points and cyanogen bromide cleavage patterns. The conversion of PROT' to PROT, which is regulated by the unlinked locus Mep, probably involves proteolysis alone, since 1) both PROT' and PROT appear to be glycosylated to the same extent; and 2) cyanogen bromide cleavage of 3H-leucine-labeled PROT' and PROT suggests a leucine-rich amino acid tract is removed from PROT' in the formation of PROT.
Alan L. Kriz and Drew Schwartz
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}