Reports differ as to the stability of Mu-induced mutants. Ben Burr (pers. comm.) has found a high reversion rate of Mu-induced sh1 mutants. Freeling has reported that some Mu-induced Adh1 mutants are unstable. In the previous report, we described a putative Mu-induced wx mutant that appears to be highly unstable.
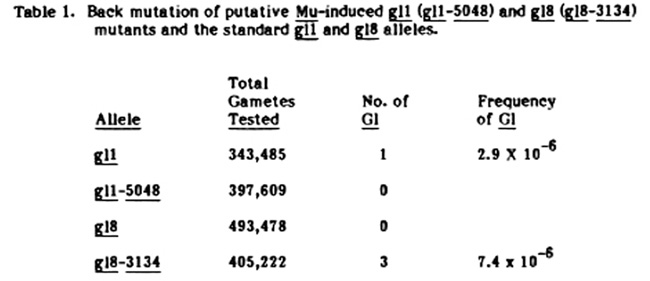
In the last few years, we have tested three putative Mu-induced mutants for their reversion rates to their respective wildtype alleles. Putative Mu-induced gl1 and gl8 mutants and standard gl1 and gl8 alleles were transferred to a y1 wx background. Each of these glossy white endosperm waxy stocks were grown in isolation and allowed to self-pollinate. The resulting white waxy seeds were grown in the seedling bench, and the frequency of non-glossy seedlings was determined. The non-glossy seedlings were transplanted to pots and the pollen of mature plants checked to confirm that they were homozygous waxy and, when possible, either self-pollinated or outcrossed to a y1 wx stock. Any plant that did not test homozygous for waxy (and white endosperm when this test was possible) was not counted as a reversion. The results of these tests are given in Table 1.
There is no indication that the Mu-induced gl1 mutant has a higher mutation rate than the standard allele. The gl8-3134 allele may have a slightly higher reversion frequency than the standard gl8 allele. A larger population of gl1-5048 will have to be grown to determine if this mutant ever reverts to its Gl allele.
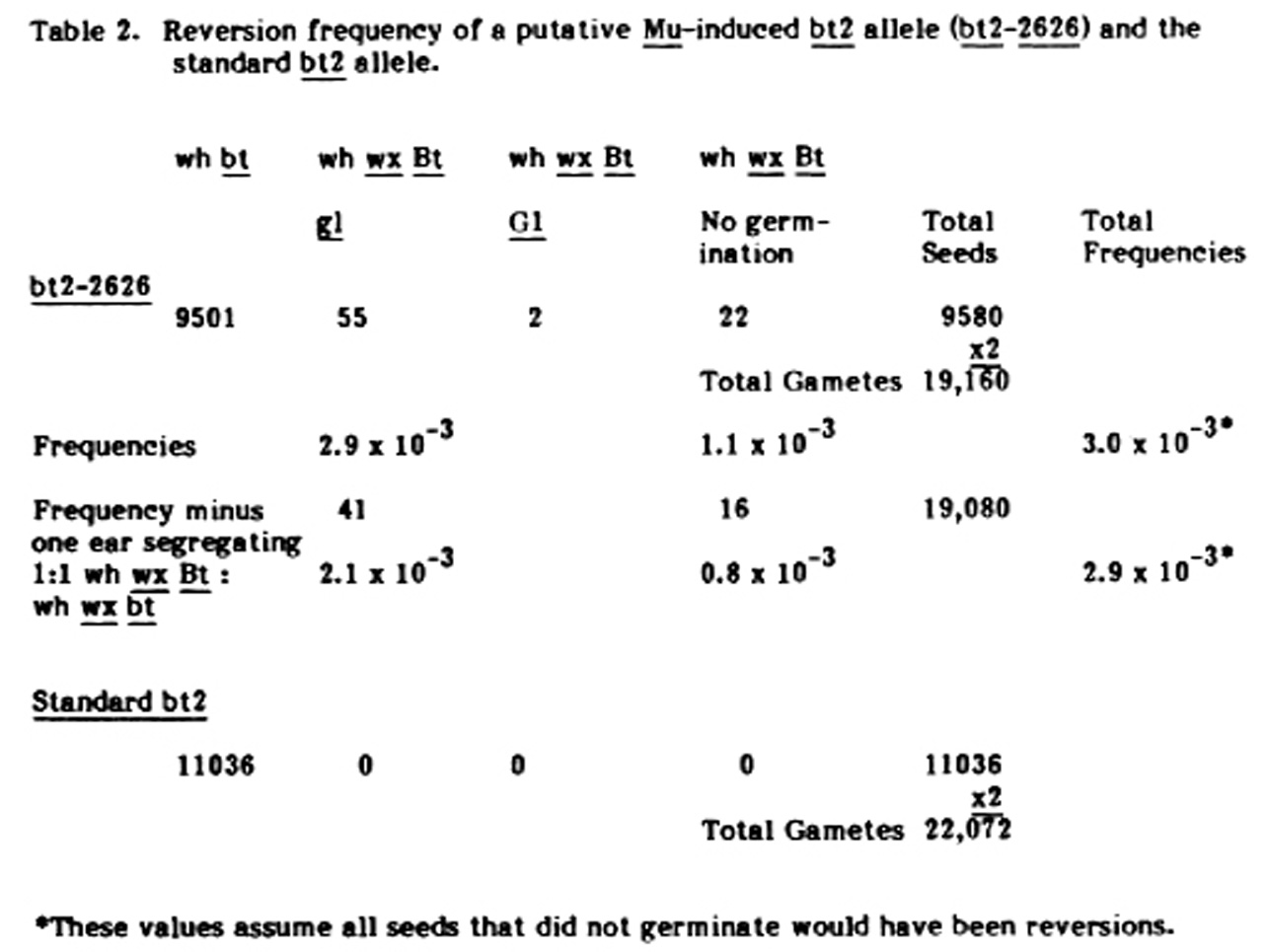
A putative bt2 Mu-induced allele (bt2-2626) and the standard bt2 allele were transferred into a y1 wx gl1 background. Once these stocks were produced they were either self-pollianted or sib-crossed and the resulting seeds were scored for white waxy non-brittle seeds. These putative reversion seeds were seedling tested to determine their glossy constitution. The results of these tests are given in Table 2. It is obvious that the bt2-2626 allele is very unstable compared to the standard allele. The reversion frequency of this allele is between 2-3 x 10-3. One selfed ear segregated 20 white, brittle:20 white, waxy, nonbrittle seeds. Since a white, waxy brittle seed was planted, the only way a 1:1 ratio would result is if the bt2-2626 allele reverted in a progenitor cell that gave rise to all of the germinal tissue of the ear or the tassel. Thus the gametes of one inflorescence would be heterozygous for the Bt revertant allele and the other would be homozygous for the mutant bt2-2626 allele. All other ears with Bt seeds had only one or two or, at the most, 3 Bt seeds. Because selfs and sibs -were involved and also because seed set was irregular on most ears, sectors could not be recognized. The high frequency of single reversions and possible small sectors with reversions suggest that reversion (back mutation?) occurs late in somatic development or possibly in meiosis. This is the same time that Mutator seems to be active in producing forward mutations. There also have been some indications that Mu may also be active in early developmental stages. Thus, it would appear that both forward and reverse mutations occur at the same time in ontogeny in Mutator stocks.
The information obtained to date on the stability of Mu-induced mutants suggests that they are variable in this regard. Some appear to be similar to the standard allele, while others revert at a much higher rate than their standard counterparts, and still others appear to be extremely stable. About 40 percent of all Mu-induced mutants are known to be somatically unstable (i.e., mutable). It could be that these also are germinally unstable. This relationship between somatic and germinal instability, if any, as yet has not been established for Mu. Somatic instability in the endosperm is expressed as very small round dots. Such a pattern would not be discernible in the brittle endosperm. It would also be difficult to recognize instability in glossy plants because of the small size of the revertant areas normally seen in unstable Mu-induced seedling mutants. This past summer we tested several Mu-induced a1 mutable mutants to determine if there is a relationship between the rate of somatic reversions and germinal reversions. The results are yet to be analyzed. The stable gl1-5048 allele could possibly be a deletion mutant. Such a mutant might be expected if the Mu1 DNA insert transposed into, or near, the gl1 locus and then transposed again, taking with it some or all of the structural gene DNA and producing a deletion. We will have to wait on the molecular geneticist for answers to some of these problems with respect to the stability of Mu-induced mutants.
Table 1. Back mutation of putative Mu-induced gl1 (gl1-5048) and gl8 (gl8-3134) mutants and the standard gl1 and gl8 alleles.
Table 2. Reversion frequency of a putative Mu-induced bt2 allele (bt2-2626) and the standard bt2 allele.
Donald S. Robertson
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}