Solubilization of thylakoid membranes under mild conditions prevents the complete dissociation of chlorophyll from protein and results in the appearance of a series of green bands (chl-proteins) during SDS or LDS PAGE. In 1977 Hayden and Hopkins (Can. J. Bot. 55:2525-2529) described a minor chla-protein from maize thylakoids which they assigned to the photosystem II reaction center. As techniques for preserving the association between chlorophylls and their associated apoproteins progressed, at least two minor chla-proteins were shown to occupy the gel region described by Hayden and Hopkins. Differential functions have not been assigned to these two chl-proteins, and their precise relationship to the primary photochemistry of photosystem II is unclear.
We investigated the association of these complexes with photosystem II by comparing polypeptide and chl-protein profiles from wild type and hcf*-3 chloroplasts under conditions of mild solubilization (4 C, 10 minutes, at pH 7.8) and electrophoresis (2 C, 10-17% gradient gels, in the dark). The nuclear hcf*-3 maize mutant is known to specifically lack photosystem II reaction centers in both mesophyll and bundle sheath chloroplasts (Leto et al., Plant Physiol. 69:1450-1458; MNL 54:117-118), and thus provides a reference for determining structural association of polypeptides with photosystem II. We also examined the chl-protein profile of purified photosystem II particles (Mullet and Arntzen, Biochim. Biophys. Acta 589:100-117) to determine which chl-proteins in the "CPa" region were essential for photosystem II activity. Minor chl-proteins in the "CPa" region were detected by activating chl fluorescence with a longwave ultraviolet source (Machold et al., Carlsberg Res. Commun. 44:235-254).
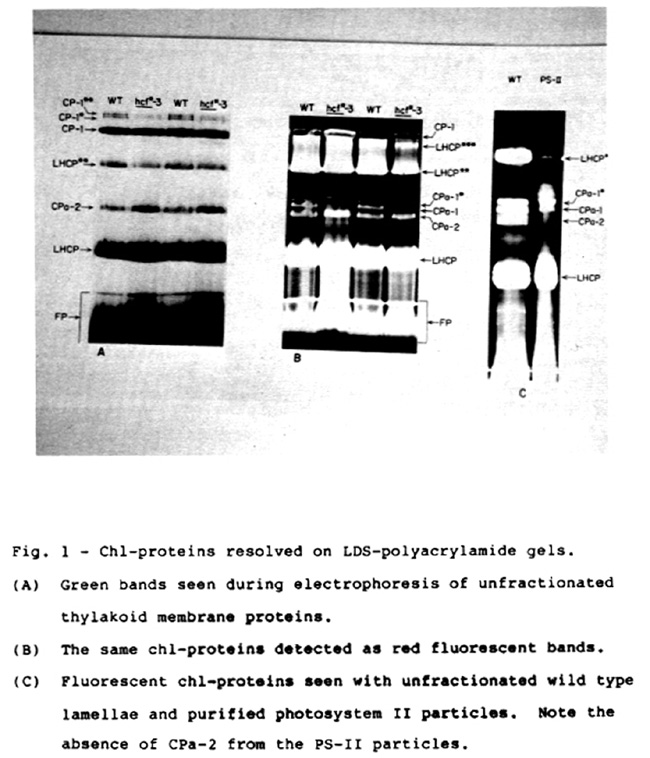
As shown in Figure 1A, several chl-proteins were resolved during LDS-PAGE at 4 C. The band labeled CPa-2 corresponds to the minor chla-protein originally described by Hayden and Hopkins. Illumination of the gel with ultraviolet light revealed that the "CPa" region contained three distinct bands, two of which, CPa-1 and CPa-1*, were clearly missing from hcf*-3 thylakoids (Fig. 1B). In contrast, the chl-protein profile from purified photosystem II particles contained both CPa-1 and CPa-1*, but lacked CPa-2 (Fig. 1C). These observations suggest that CPa-1 and CPa-1* are closely associated with photosystem II, while CPa-2 is not necessary for photosystem II activity. Further work with digitonin/triton fractionated lamellae indicates that CPa-2 is enriched in stroma lamellae and thus may be preferentially associated with photosystem I.
CPa-1 and CPa-2 were purified by preparative electrophoresis, the pigment extracted, and the apparent molecular weight of the constituent apoproteins determined by LDS-PAGE. The apoproteins obtained in this way were compared with the corresponding authentic chl-proteins by proteolytic mapping to establish identity (data not shown). This analysis assigns apparent molecular masses of 48kD and 42kD to the apoproteins of CPa-1 and CPa-2 respectively. By similar analysis CPa-1* was shown to be an electrophoretic variant of CPa-1. The general characteristics of CPa-1 make it a leading candidate for the photochemically active reaction center protein of the photosystem II complex.
Fig. 1 - Chl-proteins resolved on LDS-polyacrylamide gels.
Kenneth J. Leto and Roslyn Young
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}