The constitutive heterochromatin (or repetitive DNA sequences) discernible by knob-heterochromatin, C- and Q-bands in Zea has been the subject of speculation for many years. Much variation in shape, size and position of C-heterochromatin is found in both maize and teosinte. The evolution of repeated sequences by unequal crossing over has been indicated by Smith (Science 191:528-538, 1976), showing the possibility of increase or decrease of tandem repeats of repetitive DNA.
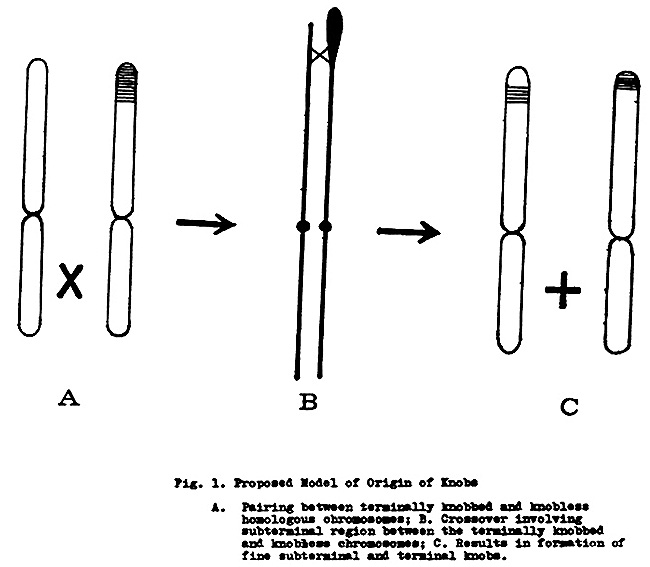
We have studied the distribution of C-heterochromatin in different races of maize and teosinte. Our observation of Imbricado, which has subterminal bands, and Confite Morocho, which has terminal bands on the 6th and 7th chromosomes (though both are from the high land of the Andes), suggests a model of crossing over between large terminal banded types and bandless types (Fig. 1) giving rise to two resultant chromosome types, one type having fine subterminal and the other having fine terminal C-heterochromatin.
Suppose that the proposed bandless type of parent had a short repetition of DNA not manifested as a C-band or knob, homologous to a DNA repetition in the band on the terminally banded chromosome type. Chance recombination could have occurred in low frequency at this heterochromatic region and after many generations of unequal crossing over at meiosis, given rise to fine subterminal and terminal bands. The structure of condensation of heterochromatin blocks, permitting crossing over only at the distal region of the band, with the main heterochromatic block looping out at the time of pairing, means that fine subterminal and terminal bands could result. This would explain the variable size of bands in different races as products of varying degrees of crossing over of the heterochromatin. Also Miklos and Nankivell's (Chromosoma 56:143-167, 1976) observation could explain why knobs/bands are inherited in Mendelian fashion and recombination does not normally occur within a knob to yield subterminal and terminal knobs in an observable frequency.
Fig. 1. Proposed Model of Origin of Knobs
Andy Pereira and J. K. S. Sachan
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}