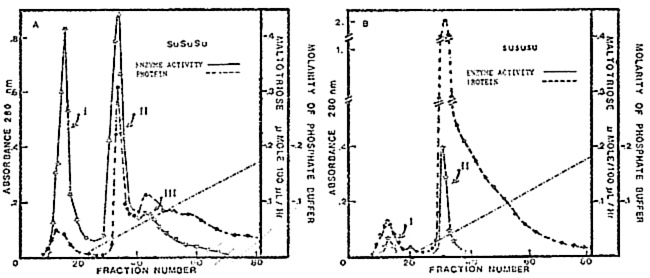
In an earlier study (Plant Physiol., 1984), we have shown that the debranching enzyme (DBE) activity from nonmutant endosperms can be separated into three peaks (1, 11, and 111) on a hydroxyapatite column (Fig. A). The extract of sugary endosperms lacks one (peak III) of these peaks of activity while the other two (I and II) have much reduced activity, particularly peak I (Fig. B).
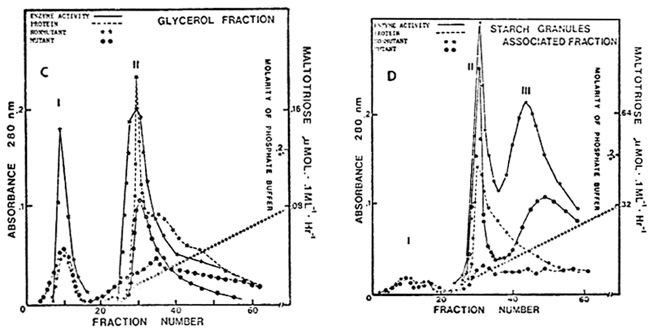
A nonaqueous procedure using glycerol as an extracting solvent (Plant Physiol. 67:518-524, 1981) was employed for the isolation of starch granules and the localization of DBE activity from nonmutant and sugary endosperms. The endosperms were initially extracted with anhydrous glycerol in a Virtis blender, followed with three glycerol washes; the glycerol-washed supernatants were combined and considered as glycerol extract (cytosol fraction); the starch granule pellets were finally washed with 0.1 M citrate buffer, pH 6.0 four times. The combined buffer-washed fraction was designated as starch granule-associated fraction (amyloplast enzyme). The extract and buffer-washed fraction from starch granules were further subjected to ammonium sulfate fractionation and DEAEcellulose and hydroxyapatite column chromatography as previously reported. In the nonmutant inbred W64A, two soluble peaks (peak I and II) and two starch granule-associated peaks (peak II and III) of DEB activity were found in the glycerol extract (cytosol enzyme) and the buffer-washed fraction from starch granules (amyloplast enzyme) respectively, indicating that peak I is mainly present in the cytosol compartment whereas peak III enzyme is essentially localized in amyloplasts; peak II is apparently distributed in both compartments. In the sugary mutant, only peak II enzyme showed in the glycerol extract while peak II and III both appeared in the buffer-washed fraction.
A comparison of the level of DBE activity in peaks I, II and III in the glycerol extract and the buffer-washed fraction from starch granules, from nonmutant and sugary mutant endosperms, indicates the deficiency of peak I (cytosol compartment enzyme), and a much reduced activity of peak III (amyloplast enzyme) in the sugary mutant. A detailed analysis of the elution patterns of peaks I and III, which were prepared by two different extraction procedures (aqueous and nonaqueous), on hydroxyapatite columns implies that there is a reciprocal relationship in enzyme activity between peaks I and III in that for Su/Su/Su endosperms column chromatography of buffer extracts shows substantial peak I and lower peak III activity (Fig. A). However, there is very low peak III activity in the glycerol washings, and the buffer wash of the glycerol-extracted starch has little peak I activity and higher peak III activity than the buffer extracts of the endosperms (Figs. C and D). We can only speculate at the present time as to the reason for the differences in peaks I and III activity with two different extraction techniques. It is possible that Su and su code for a polypeptide which as a homodimer constitutes peak III only in the absence of cytosol components (nonaqueous extraction), but can combine with the product, present in the cytosol compartment, of another locus to form a heteromeric enzyme (peak I) at the expense of peak III when the endosperms were buffer-extracted. The study on the characteristics of these three peaks is in progress.
David Pan
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}