As pointed out in previous articles (MNL 52:37, 1978, Magoja, Com. Direc. Inv. U.N.L.Z. 2 (3):10, 1980), the fertility of hybrids between perennial teosinte and Gaspe is relatively high. The triploid hybrids obtained between perennial teosinte and other maize varieties are generally highly sterile. This phenomenon has been noted by some other authors as well as ourselves. It is well known that triploidy per se conditions high sterility; fertility analyses of different hybrids between perennial teosinte and maize indicate that the grade of genetic affinity of their progenitors has a considerable influence on fertility. There is no doubt, therefore, that the chromosome imbalance of these hybrids does not constitute the only cause of sterility. Consequently, the appropriate selection of maize genotypes, which combined with perennial teosinte will produce hybrids with high fertility, must lead to the attainment of fertile hybrids.
Intending to measure fertility of perennial teosinte-Gaspe hybrids, pollen viability was evaluated in F1 and F2 populations. An F1 population was studied which was comprised of 110 plants originating from the same number of hybrid seeds, obtained by crossing perennial teosinte with Gaspe. The F2 population of 598 plants originated from the seed obtained on F1 plants, by way of free pollination between themselves. The pollen of each individual plant was harvested during the 1982/1983 summer in both populations. The pollen was stained with I2-KI sol. mixed with an equal volume of lactic acid, and was observed with a microscope. A count was carried out over approximately 200 grains of pollen of each plant. Only the pollen grains that colored totally with the stain were considered fertile.
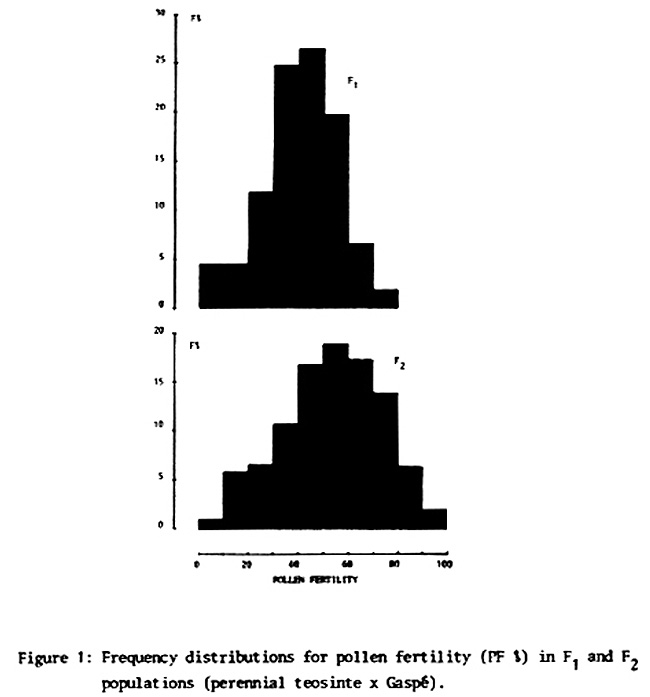
In Figure 1 the frequency distribution of fertility for F1 and F2 can be seen. Although in F2 the variability observed can be explained on the basis of different genetic combinations that are produced, the high variability of F1 cannot be attributed to environmental causes. It is evident that the variability of fertility of the F1 is due to expression of different F1 genotypes, perhaps brought about by the different gametes contributed by perennial teosinte.
In order to have an estimate of the environmental component of the total F1 variability, numerous plants were cloned. The clones of each F1 plant (equal genotype) showed that the variability of conditioned fertility through environment is approximately 33%. In other words, variability of fertility among F1 plants is due in 67% to genetic causes.
The results obtained in F1 and F2 populations are summed up in Table 1.
It can be noted that the average fertility increases in successive generations of hybrid progenies of perennial teosinte and maize. While we do not present results, fertility increases in the F3. In that the hybrid progenies were obtained without any selection pressure, it is evident that there is a natural preferential selection of better balanced genotypes and chromosomal endowment, which conditions greater fertility.
Comparing the fertility of hybrids between perennial teosinte and Gaspe with other hybrids which we have obtained employing other maize, we were able to deduce that the former are relatively fertile and that perennial teosinte and Gaspe produce good genetic combinations for fertility. While the fertility of F1 of the hybrids between teosinte and Gaspe is an average of 41.2%, other hybrids obtained do not reach 15%. In F2 progenies, the hybrids between teosinte and Gaspe have 53.4% mean fertility, and other hybrids only 28% average fertility.
The progenies of hybrids between teosinte and Gaspe can yield individuals nearly totally fertile in generation F2. Fertility of hybrids between perennial teosinte and Gaspe approaches the much higher fertility of hybrids between maize and the other teosinte species.
Ida Graciela Palacios and Jorge Luis Magoja
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}