A majority of the studies dealing with pachytene cytology of maize characterize the genome in terms of relative length. In the present report, an attempt was made to analyze the pachytene karyomorphology of ten cultivars of maize, mostly of Indian origin, in absolute terms, and the results were compared with those of Longley (J. Agr. Res., 59:475-490) and Maguire (Cytologia, 27:248-257). The patterns of length variability were examined by regression analysis of pooled data. In addition, observations were made regarding other quantitative features of meiosis.
The course of meiosis in all the cultivars is uniformly regular, except for minor quantitative variations in chiasma frequency and the nature of bivalents. The total lengths of the haploid complements range from 860.09 u (in Adecuba) to 512.81 u (in Pira Cundinamarca).
Of the ten cultivars analyzed in the present investigation, three (Stowells Evergreen, Kalimpong Local and Sonada Local) have knobless pachytene chromosomes. The knob number in other cultivars ranges up to a maximum of 11 in the case of Pira Cundinamarca. The knob constitution in some of the cultivars was found to be variable from plant to plant. This is especially true of composites such as Vijay and Diara, and synthetics such as Synthetic B19. The various methods of hybridity, selection and bulking may have contributed to the observed variation. Adecuba, Darjeeling Market, Diara and Synthetic B19 may be considered as low knob numbered, whereas the rest are medium knob numbered, excludinq the highly numbered Pira Cundinamarca and the knobless ones. The IKL-1 is the most frequently encountered knob in the present investigation.
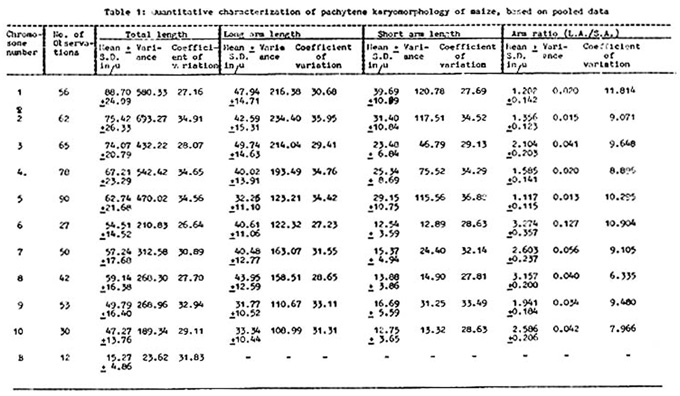
Absolute measurements are enormously variable. Arm ratios are more reliable than absolute lengths for chromosome identification. The pooled data of the absolute measurements of eight cultivars are presented in Table 1 along with other relevant statistical parameters.
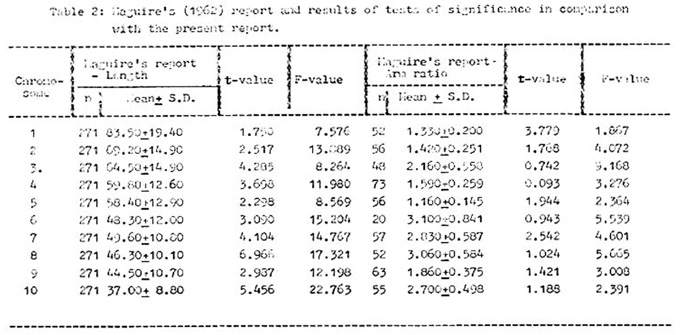
Significance tests such as t- and F-tests were conducted to compare the means and variances of total length and arm ratios from the present report with those of Maguire's data (op. cit.), the results of which are given in Table 2, along with the latter report.
The mean total lengths of chromosomes 3, 4, 7, 8 and 10 from the present investigation are significantly different, whereas those for chromosomes 6 and 9 are significant at the 50% level but not at the 1% level. However, the means for chromosomes 1, 2 and 5 do not show significant differences. It is to be noted that chromosomes 1, 2 and 5 all have mean arm ratios less than 1.356 (nearly median chromosomes). However, the estimated variances of lengths show significant differences.
With respect to arm ratios, t-values for most of the chromosomes are insignificant. For chromosome 1, the t-value is highly significant and for chromosome 7, the same is significant at the 2.5% level, but not at the 1% level. Estimated variances of arm ratios are highly significant except for chromosome 1, the F-value of which is significant at the 5%, but not at the 1% level.
With respect to comparison of the present data with those of Longley, it is worth mentioning that Maguire obtained insignificant t- and F-values for all the chromosomes except for chromosome 7 (with a reciprocal translocation) during the comparative study of total length. Her mean arm ratios are quite close to those of Longley. However, it is not possible to carry out F-tests individually for each arm ratio, in view of the nature of the data presented by Longley. This indicates that the total lengths of chromosomes in the present data are considerably different from those of Longley, whereas most of the arm ratios are remarkably similar.
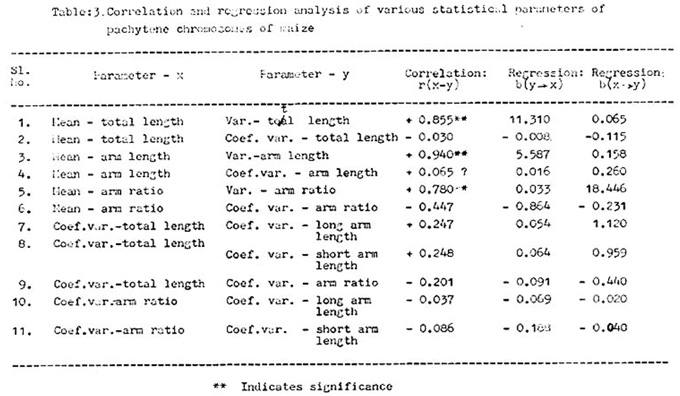
Correlation and regression analyses were conducted to analyze the relation of various parameters and the results are presented in Table 3.
Further extending the statistical analysis, it was observed that the longer the chromosome, the greater the variability in length, as was evident from a highly significant positive correlation between mean and variance of total length, and also from the fact that coefficient of variation of total length does not vary much from chromosome to chromosome. This corroborates the findings of Maguire that variability may tend to be distributed uniformly per unit length. This is also true individually for arm length and arm ratios, in view of the highly significant positive correlation coefficients.
Short arms are not consistently either more or less variable in length than the long arms, if the differences in their averages are taken into account. This is supported by the finding that coefficients of variation of arm length do not vary much between short arm and long arm. Thus the conclusion of Maguire (op. cit.) that the short arm is more variable than the long arm in a chromosome, does not coincide with that from the present data.
It appears that relative variability (coef. var.) of arm ratio has no significant influence on relative variability in total length. In fact, there is a slight negative correlation observed between these two characters. This is also true if the variabilities in long arm length and short arm length are separately examined.
The possible reason for the deviation of the results of the present report from those of Maguire and of Longley becomes quite intelligible if the nature of the materials and method of analysis are taken into account. Longley's data on length measurements represent pooled data of all the chromosomes in 28 cells from 14 Indian varieties. He also gave mean total length, arm lengths and arm ratios for 33 Indian varieties and 41 Mexican varieties. Maguire's data represent length measurements from all the chromosomes of 271 cells. For arm ratios, different numbers are measured for each chromosome. The materials included 6th and 7th backcross generations (to maize) of a maize-Tripsacum hybrid. The maize background included L289 and CC5, and a few had a reciprocal translocation. She reported a significantly different F-value for chromosome 7, which was the Zea-Tripsacum interchange chromosome. In these two reports, the same number of observations were not made for total lengths and arm ratios. The present analysis is based on pooled data from 8 cultivars (mostly Indian). The materials included are relatively more heterogeneous than those of Maguire, but less heterogeneous than those of Longley. Another factor of difference is that the same number of observations has been made, both for total lengths and arm ratios. These factors might have been responsible for the discrepancy in the present data on total lengths, but are more or less in complete agreement with respect to arm ratios, when compared with the previous records.
Based on the present data, the following conclusions can be made:
1. Variability of total length, arm length and arm ratio may be uniform per unit length.
2. Differences in the variabilities of long and short arms in a chromosome are not considerably apparent.
3. The relative variability of arm ratio has no significant influence on relative variability of total lengths.
The present investigation was carried out under the supervision of Prof. A. K. Sharma of the Department of Botany, University of Calcutta, Calcutta.
J. S. P. Sarma
Return to the MNL 57 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}