One widely used maize test system for mutagenesis utilizes the yellow-green-2 (Yg2) locus. In several studies, dry or soaked kernels heterozygous for this locus have been treated with chemical or physical agents, and the resultant seedlings were classified for the frequency of sectors expressing the recessive (yellow-green) phenotype (Latterell and Steffensen, 1962, Am. J. Bot. 49:478; Smith and Rossi, 1966, Rad. Res. 28:302; Conger and Carabia, 1977, Mut. Res. 46: 285; Fujii, 1980, Jap. J. Genet. 55:241; Plewa, 1982, pp. 411-419 in Environmental Mutagens and Carcinogens, ed. Sugimura, Kondo, and Takabe; and several other studies). For studies of the mutagenic activity of physical and chemical agents, the fourth and fifth leaves are typically classified. In these studies, it is usually acknowledged that the mutant sectors can originate in two distinct ways: (1) by mutation of the dominant allele at the yellow-green locus or (2) by a deficiency of the dominant allele of this locus. Because plants heterozygous for large terminal deficiencies of this chromosome, including much of the short arm, are known to be viable (McClintock, 1943, Carnegie Inst. Wash. Yearbook 42:148), and plants homozygous for smaller deficiencies including this locus are also viable (McClintock, 1944, Genetics 29:478), it is likely that a significant proportion of the sectors of yellow-green phenotype are due to deletions of the Yg2 locus. Furthermore, loss of an entire chromosome 9 in this test system would be tolerated because plants monosomic for chromosome 9 survive to maturity and are quite vigorous (Weber, 1973; Theor. Appl. Genet. 43:167).
Clearly, it is important to be able to distinguish between these two types of mutational events (terminal deficiencies including the Yg2 locus and gene mutations at the Yg2 locus) in mutagenesis studies. In this paper, I propose a modification of this test system which can be used to distinguish between these two types of events. It is also important to recognize that part of the sectors of yellow-green phenotype could also be due to events which do not involve the Yg2 locus. For this reason, an additional control series should be included in these tests. This neglected important control series will be discussed at the end of this report.
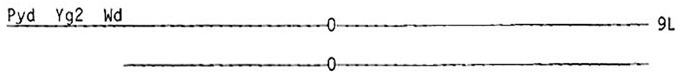
McClintock (1944) generated a large series of terminal deficiencies involving the short arm of chromosome 9 in maize, and she determined that plants homozygous for a deficiency including the knob and half of the most distal chromomere were albino. Thus, a factor which is necessary for chlorophyll synthesis lies distal to this breakpoint very close to the end of the short arm of chromosome 9. She named this factor the Wd factor (for white deficiency). A chromosome with this terminal deficiency is termed a wd chromosome. She also found that plants homozygous for an even smaller terminal deficiency of the short arm of chromosome 9 (deficient for the region distal to the first chromomere) were pale yellow. Thus, another factor, Pyd (for pale-yellow deficiency), is located distal to the wd locus, and the pale-yellow deficiency uncovered this locus. She also determined that the yg2 locus was uncovered by the wd deficiency but not by the pyd deficiency; thus, the yg2 locus lies between the wd and pyd breakpoints and all three factors are extremely close to each other or near the end of the short arm of chromosome nine distal to the terminal half of the most terminal chromomere.
The wd/Yg2 test system: Maize plants can be easily generated which carry one normal chromosome 9 containing the dominant alleles of the Yg2, Pyd, and Wd loci and a chromosome 9 carrying the wd deficiency (which deletes all three loci):
This plant is hemizygous for all three loci. Kernels of this genotype could then be exposed to chemical or physical agents. If a forward mutation occurred at the Pyd, Yg2, or Wd locus, a pale-yellow, yellow-green, or albino sector on an otherwise green leaf would be produced. However, if a break occurred proximal to the Wd locus in the normal chromosome, an albino phenotype would be produced because the Wd locus is deleted. Mutations at the Wd locus and deficiencies including the Wd locus would both be expressed as the same mutant phenotype. However, it is not unreasonable to assume that the forward mutation frequency determined at this locus would be similar to the mutation frequencies present at the other two test loci, and this estimated mutation frequency could then be subtracted from the total frequency of albino sectors. In this way, it would be possible to correct for forward mutation at the Wd locus. Albino sectors generated by loss or mutation of the Wd locus should be easily distinguishable from yellow-green and pale-yellow sectors generated by mutation at those loci; however, it is possible that pale-yellow and yellow-green sectors cannot be distinguished from each other.
It is important to note that the Pyd, Yg2 and Wd loci are extremely close to each other near the tip of chromosome 9, and it would be an extremely rare event where a break occurred distal to the Wd locus but proximal to the Yg2 or Pyd locus to generate a sector of yellow-green or pale-yellow. For this reason, expression of the Yg2 or Pyd recessive phenotypes would almost invariably be due to gene mutation at the respective loci. On the other hand, any breakpoint proximal to the Wd locus on the chromosome bearing the dominant alleles will generate an albino sector.
Seed of the above type would be treated and the frequency of albino, pale-yellow, and yellow-green sectors determined on the fourth or fifth leaves. The forward mutation frequencies at the Pyd and Yg2 loci are the number of yellow-green or pale-yellow sectors divided by the number of progenitor cells present in the kernel at the time of treatment (minus the frequency in the untreated control). The frequency of loss of the Wd locus is the frequency of albino sectors minus the mean frequency of yellow-green and pale-yellow sectors.
Generation of the test plants: The plants used in the above test system can easily be generated in very large numbers by making the cross described below. Plants homozygous for the Wd deficiency, homozygous for the C locus and all other dominant seed color factors, and also containing the Wd ring-chromosome (which contains the dominant alleles of the Wd, Pyd, and Yg2 loci and also C-I which is dominant to C and inhibits anthocyanin synthesis could be crossed by nearly any line of corn as either the male or female parent. The dominant alleles of the Wd, Pyd and Yg2 loci will be present in any true-breeding, non-chlorotic line of corn, including commercial varieties, and all factors necessary for kernel pigmentation are contributed by the ring-containing parent. Two types of progeny are produced, those containing the ring chromosome and those lacking the ring. The ring-containing progeny can be easily identified because the C-I locus is present on the ring, and kernels containing the ring have colorless endosperm (in tissue where the ring chromosome is present) with sectors of colored tissue (where the ring is lost). Kernels lacking the ring chromosome are colored because the C-I locus is not present, and these represent the vast majority of the progeny because small ring chromosomes are unstable, are frequently lost, and are present in only a small proportion of the cells of the organism (McClintock, 1938, Genetics 23:315). The latter kernel type (which lacks the Wd ring) would be used in mutagenesis studies described above. Kernels of this type are being generated and will be used in mutagenic studies in the near future. wd/wd ring-containing germplasm is available from several laboratories, including ours, and plants of this type can be increased simply by selfing or sibbing. The ring-containing wd/wd kernels from such a self can be readily identified because they have endosperm which is colorless and also contain purple sectors.
A neglected important control: Another shortcoming of tests using plants heterozygous for the Yg2 locus is that it is not known if some of the mutant sectors are due to events other than mutation or loss of the Yg2 locus. It is not unlikely that dominant mutations at other loci can occur which produce a mutant phenotype indistinguishable from the Yg2 phenotype. Also, in my work with monosomics, I have found several plants in which half of each leaf is lighter green than the other half, and this altered phenotype is associated with monosomy. In these cases, the marker mutations utilized in selection of monosomic types (g and j) are not associated with the altered phenotype (Weber, 1973, Theor. Appl. Genet. 43:167; 1982, pp. 79-83 in Maize for Biological Research, W. F. Sheridan, ed.). Clearly, hemizygosity of certain unknown chromosomal segments can result in a lighter leaf phenotype, and this phenotype is not associated with the Yg2 locus. For this reason, an important control for this type of work would be to treat homozygous dominant (Yg2/Yg2) plants with the agent being tested to determine if the agent causes mutant sectors which are not associated with loss or mutation of the Yg2 locus. If such sectors are found, the frequency of such sectors minus the square of the mutant sector frequency in the experimental group (to correct for types where mutations took place at both Yg2 loci) would need to be subtracted from the frequency in the experimental group to correct for events which do not involve the Yg2 locus.
In fact, Neuffer (1966, Genetics 53:541) noted that this case was indeed found in plants grown from EMS-treated seed. Although no data were given, he mentioned that the frequency of mutant sectors on leaves of Yg2/Yg2 plants was higher than
the square of the frequency found in Yg2/yg2 plants. It is important that these control plants be included in future tests. (Supported in part by DOE Contract 79EV02121.)
David Weber
Return to the MNL 57 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}