In the last year, several papers have appeared on the structure of HC-toxin, a phytotoxin produced by the maize pathogen Helminthosporium (perfect stage Cochliobolus) carbonum race 1. This toxin has the same host range as does the fungus and is thus "host-specific." Both H. carbonum race 1 and HC-toxin selectively affect maize that is homozygous recessive at the nuclear Hm locus, located on the long arm of the first chromosome.
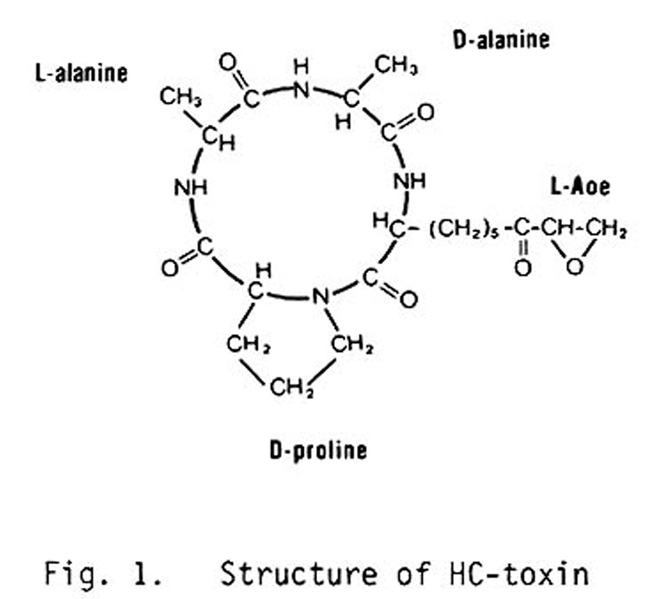
The structure of HC-toxin, as established and confirmed by four different laboratories, is a cyclic tetrapeptide, cyclo(L-Aoe-D-prolyl-L-alanyl-D-alanyl) (Fig. 1), where Aoe stands for 2-amino-8-oxo-9,10-epoxidecanoic acid (J. M. Liesch et al., 1982, Tetrahedron 38:45-8; J. D. Walton et al., 1982, Biochem. Biophys. Res. Comm. 107:785-94; M. R. Pope et al., 1982, Phytopathology 72:941; M. Kawai, D. H. Rich, J. D. Walton, B. W. Gibson, in preparation). Walton et al. (1982) purified HC-toxin from culture filtrates of the fungus by solvent extraction, gel filtration, and HPLC. The MW and elemental composition were determined by high resolution fast atom bombardment (FAB) mass spectrometry. The amino acid composition was determined with an automatic amino acid analyzer and by 1H and 13C NMR, and their sequence by GC-MS of the derivatized products of a partial acid hydrolyzate. This last step was especially important because cyclic peptides are notoriously difficult to sequence by other techniques. Our original conclusion that HC-toxin contained two L-alanine residues has been re-examined in light of NMR studies on the conformation of HC-toxin by Daniel Rich at the University of Wisconsin, suggesting that the second alanine has the D configuration, and we have now established by two criteria (HPLC and automatic amino acid analysis of D- and L-amino acid oxidase digests of an acid hydrolyzate of HC-toxin) that this is, in fact, the case (M. Kawai et al., in preparation).
HC-toxin has elemental composition C21H32N4O6 and MW 436. It is unusual in being both chloroform and water soluble. It can be isolated from culture filtrates of H. carbonum race 1 in up to 10 mg/l quantities. It inhibits root growth half maximally at between 0.2 and 1.0 ug/ml; resistant maize is affected at approximately 100 times higher concentration.
The unusual epoxide-containing alpha-amino acid Aoe has been found previously in two other fungal cyclic tetrapeptides, the phytotoxin Cyl-2, cyclo(L-Aoe-D-0-methyltyrosine-L-isoleucine-L-pipecolic acid), and the cytostatic agent chlamydocin, cyclo(L-Aoe-alpha-aminoisobutyryl-L-phenylalanyl-D-Drolyl). Recent studies by Daniel Rich have shown that the peptide backbones of chlamydocin and HC-toxin have the same conformation in chloroform. Both chlamydocin and HC-toxin (J. D. Walton and E. D. Earle, 1983, Physiol. Plant Path., in press) require an intact epoxide for activity. Based on several criteria, it seems likely to us that chlamydocin and HC-toxin have the same site of action, which is still unknown. Chlamydocin is much less host-specific than HC-toxin; although HC-toxin-sensitive roots (genotype hm hm) are equally sensitive to HC-toxin and to chlamydocin, HC-toxin-resistant roots (genotype Hm Hm or Hm hm) are much more sensitive to chlamydocin that to HC-toxin (unpublished observations). Mammalian cells in culture are sensitive to both chlamydocin and HC-toxin at less than 1.0 ng/ml (unpublished observations).
Fig. 1. Structure of HC-toxin
Jonathan D. Walton and Elizabeth D. Earle
Return to the MNL 57 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}