Considerable confusion surrounds the fate of the cells produced by the first few divisions of the zygote in maize. It is generally agreed that the first division after fertilization is transverse (note, however, the perplexing results of Bird and Neuffer, in this issue), and that the two products of this division probably form the embryo and the suspensor. The next division of the embryo cell is in a longitudinal plane, but its exact orientation is unclear. Traditional interpretations hold that the products of this division are destined to form the embryo and scutellum. On the other hand, Steffensen has argued that the first longitudinal division defines the midrib line, dividing the plant into left and right halves. Recent results from a clonal analysis of embryogeny indicate that neither of these views is entirely correct.
In order to conduct a clonal analysis of the relationship between the embryo and scutellum, it is necessary to use cell marker mutations that are expressed in both of these structures. For this purpose, we used the stock Wd C-I/wd C; +/R-sc. Kernels of this genotype are colorless due to the dominant color inhibitor C-I, and yield purple sectors in both the aleurone and scutellum when this gene is lost. The loss of the chromosome arm carrying C-I coincidentally exposes wd--an albino mutation that is expressed in the seedling. Thus sectors present in the scutellum can also be observed in the embryo if these two structures share the same lineage.
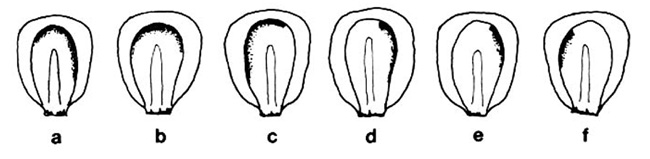
Plants of the genotype Wd C-I; r-g were pollinated by wd C, Wd 9S ring; R-sc/() and ears were x-rayed (500R, 1-mm Al, 140 kVp) 56 hrs later (note that the Wd ring chromosome is poorly transmitted through pollen, so most of the progeny from this cross are of the appropriate genotype). Histological observations indicated that specimens had undergone 1-2 longitudinal divisions by this stage. At maturity, kernels were screened for sectors in the scutellum and selected kernels were then planted to determine whether sectors extended into the seedling. Six clearly sectored seeds were observed out of a total of 815. Three of these seeds (a, b and c) gave white seedlings, and 3 gave green seedlinqs (d, e and f). In addition, one of the seeds which appeared to lack a sector in the scutellum gave rise to a white seedling with a narrow strip of green tissue along the margin of the first leaf (the apparent absence of a sector in the scutellum of this kernel is probably due to the poor expression of R-sc in the scutellum; the experiment is being repeated using R-scm). White seedlings arose from kernels in which the sector was located at the apex of the scutellum directly in line with the embryonic axis; green seedlings arose from seeds in which the sector was located along one side of the scutellum.
These data demonstrate that the scutellum and the embryo can share the same lineage after the first or second longitudinal divisions. If the lineages of these two structures were separated at this stage, sectors would never encompass both of them. It is also clear that the lineages resulting from these early divisions are not necessarily destined to form the left and right halves of the plant, otherwise sectors that encompassed 1/2 of the scutellum (Fig. 1d) would have produced half sectored plants. We have observed half sectored plants in material irradiated 6 days after pollination, when the embryo was at a globular stage. Such plants are rare, however, and it is more common to find sectors involving smaller or larger fractions of the plant body. Our observations confirm Randolph's conclusion that the early pattern of cell division during maize embryogeny is highly irregular, and provide no evidence of determinate cell lineages.
R. S. Poethig and E. H. Coe
Return to the MNL 57 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}