A large M1 from treatment of Mo17 pollen with EMS and crossing on A632 silks was planted and observed for dominant mutants. The method of treatment was that described in MNL 56:42. The purpose was to produce mutants for earliness, male sterility, short plants, and other agronomically useful traits. 7,997 kernels were planted, producing 6,425 seedlings. The M1 was notable for its high frequency of apparent genetic changes, including maternal and paternal haploids, monosomics, and dominant mutants. Among these last were 7 shredded leaves, 2 golden sheaths, 21 yellow-greens (including sectored forms, below), 4 with tillers, 3 male steriles, 5 with pubescent leaf sheaths, 35 dwarfs or very short plants, 21 with leaf lesions or leaf texture changes and a number of other miscellaneous traits.
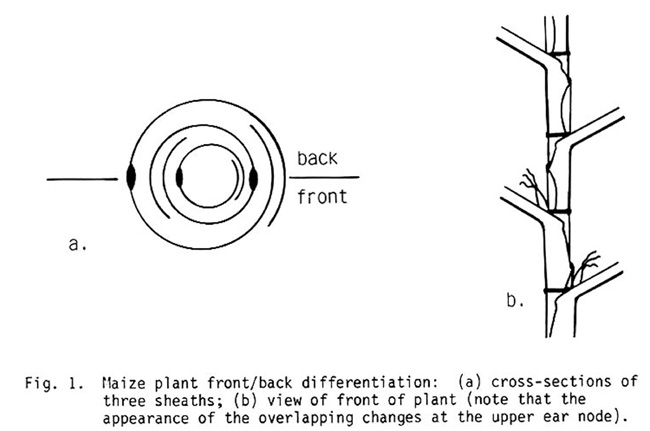
A curious pattern was observed for 54 plants--one-half the plant (divided by a plane through the leaf midribs) was phenotypically different from the other half. Lesions, striping, inter-vein narrowing, shredding, rapid aging, and other traits could be restricted to 1/2, 3/4 or 1/4 of the leaf. Often the midrib limited the affected area and sometimes there was only a narrow stripe, but for all cases, both ranks of leaves were affected, in one rank the "left" half of the leaf, in the other the "right" half. Traits restricted to one rank were not observed. We decided to use Edgar Anderson's terminology (The Corn Plant of Today, p. 17) to distinguish the two plant halves: front and back, with the front being that half of the plant with the outer sheath-overlaps (Fig. 1). About a third of the dominant mutations affecting leaves were expressed in such wide sectors (a careful count depends on observing the next generation).
It is hard to explain this unless it reflects a mutational change of one-half of a DNA double-helix in the pollen grain gametic nucleus leading to genotypic difference between the first two embryonic cells. Often these two cells would contribute equally to the plant, but one cell could contribute to only a quarter or could become the entire plant. This, however, does not coincide with L. F. Randolph's description of maize embryogeny (1936, J. Agr. Res. 53:881).
All plants were self-pollinated, if possible, and those thought to be dominant mutations at the time of pollination were crossed onto Mo17, if available, or onto another stock. Ca. 4,000 relatively full ears and 1,200 semi-sterile or poorly pollinated ears are being scored for any observable differences from untreated material. Relatively few are fully or even nearly normal, perhaps 10%. A wide range of familiar and unfamiliar mutations or aberrations are being catalogued.
Fig. 1. Maize plant front/back differentiation: (a) cross-sections of three sheaths; (b) view of front of plant (note that the appearance of the overlapping changes at the upper ear node).
Robert McK. Bird and M. G. Neuffer
Return to the MNL 57 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}