These stocks involve w15 (w*-8896) and l10, closely linked with each other and with Y vs. y. The original stocks obtained from Robertson were from selfs of Y +/y l10 and Y +/y w15. Crosses were made of the green progeny from one stock with those from the other. Ears from crosses that were between Y +/y l10 and Y +/y w15 were segregating for yellow and white endosperm. Most of the plants from the white seeds from that cross were heterozygous for w15 and for l10 in repulsion. These plants were selfed and also crossed on a Y Y normal stock. The progeny were grown in a somewhat isolated block and allowed to open pollinate, except for 54 which were self-pollinated. The results of the seedling tests for 299 ears are as follows:
All ears were segregating for yellow vs. white endosperm. There were 7 recombinants between l10 and w15, 2.34% recombination. In an earlier report (Maize Newsletter 51, 1977, p. 55), on a similar experiment using self-pollination, there were 158 that segregated l10, 155 that segregated w15, nine that had only green seedlings and three that segregated l10 and w15, a recombination value of 3.1%. Many more ears remain to be tested. There are four ears from coupling heterozygotes already identified and available for further use: one from this year's self-pollination and three from the open-pollination plot.
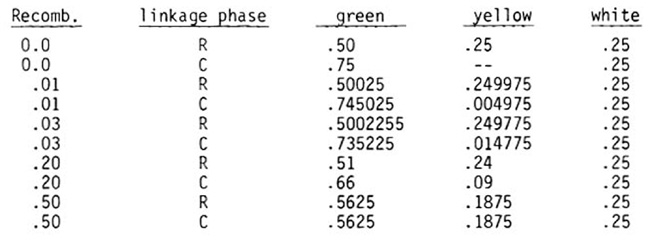
The gametic frequencies for a plant heterozygous for w15 and l10 are: p/2 ++, (1-p)/2 + w15, (1-p)/2 l10 +, and p/2 l10 w15, where p = recombination in repulsion. The frequencies of the three possible F2 phenotypic classes are: (2+p2)/4 green, (1-p2)/4 yellow, and 1/4 white. Plants that are l10 l10 w15 w15 are white.
To calculate the expected frequencies in repulsion (R), substitute for p the recombination percentage for the numbers in coupling (C), substitute 1-p for p in the expressions.
The figures for a few p values are as follows:
Note that in F2 for repulsion 50% of the chlorophyll deficient plants are yellow at 0%. recombination and 42.9% are yellow at 50% recombination. In F2 from coupling when recombination is zero, there are no yellow seedlings. At the observed 3% recombination the ratio of yellow:white seedlings is about 1:17. To have only a .01 chance of failing to obtain at least one yellow seedling, 81 plants from the white seeds would have to be grown. If Y y is not used as a marker, over 300 would need to be grown.
In this year's plot, comprised of plants from the repulsion heterozygote (y + w15/y l10 +) crossed on Y Y; at 3% recombination, about 1.5% of the plants would be coupling heterozygotes, 1.5% would be heterozygous for neither l10 or w15, and the remainder would be either l10 or w15 heterozygotes in equal numbers. Of the green survivors from selfs of the coupling heterozygotes, about 64%, of the green plants will be coupling heterozygotes, 32% homozygous green, 4% heterozygotes for w15 or l10. In the open pollination plot, the pollen mixture will be about 2 Y + +:1 y + w15:1 y l10 +. For the white seeds from open-pollinated coupling heterozygotes the ratio of yellow:white seedlings will be about 1:1. For all seedlings about 1/8 will be yellow. Most of the green plants from white seeds will be repulsion heterozygotes; a few will be from crossovers between Y vs. y and l10 or w15. About 38% of the green plants from the yellow seeds will be coupling heterozygotes. Probably the best way to obtain and maintain demonstration ears that will give these higher numbers of yellow seedlings is to grow the progeny from those open pollinated ears, and pollinate the green survivors with a mixture of pollen from progeny of Y Y x y + w15/y l10 +.
These stocks should be useful for class use-in genetics to illustrate:
1. Close linkage: Y y vs. l10 or vs. w15.
2. Balanced lethal, a repulsion heterozygote: the surviving green plants are usually heterozygotes.
3. The coupling and the repulsion heterozygotes illustrate the explanation for the true breeding of heterozygotes in many wild races of Oenothera, also the rare occurrence of types that were believed at first to be mutations.
4. The repulsion heterozygotes are useful for making crosses for linkage studies, for mapping these lethal traits.
5. Open pollination plots using this material might be useful for population theory involving closely linked factors.
Chas. R. Burnham
Return to the MNL 57 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}