It is often proposed that meiotic homologue pairing depends primarily upon prior juxtaposition of homologous centromeres and/or telomeres, with synapsis initiated first near these structures and then zipping to completion. If this indeed is the case, there is nevertheless reason to believe that there is a crucial prealignment stage which seems to occur, in some cases at least, with a frequency directly related to the extent of homologous chromosome length.
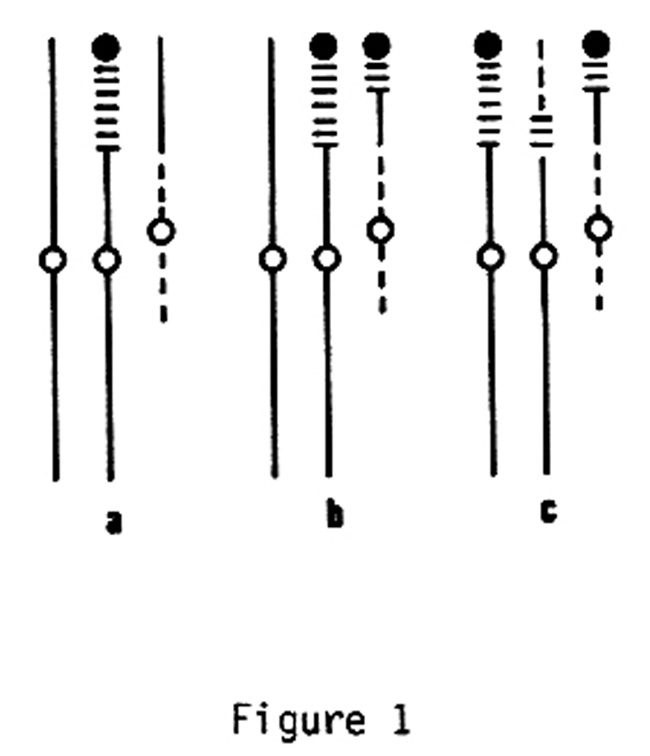
Figure 1 diagrams chromosome constitution with respect to maize chromosome 2 and a Tripsacum partial homologue in three interchange stocks (M.P. Maguire: Genetics 51:23-40; Genet. Res., Camb. 12: 21-27). Maize chromosome 2 is indicated by solid lines, the Tripsacum chromosome homoeologous portion by parallel horizontal bars, and the remainder of the Tripsacum chromosome by dotted lines; centromeres are designated by open circles, and there is a terminal knob on the Tripsacum homoeologous portion. The centromeres of maize chromosome 2 and the Tripsacum chromosome are not homologous.
Frequency of metaphase I trivalents is about 90% in types a and b, and about 54% in type c. Trivalent occurrence at metaphase I depends, in types a and c, upon homologous pairing and crossing over having occurred between the chromosome with the Tripsacum centromere and one of the two chromosomes carrying a maize centromere. In type a, with approximately twice the extent of homology available for these events (vs. type c), there is roughly twice the frequency of trivalents (vs. type c). In type b, the same total extent of homology is available for trivalent formation as in type a, but homology to the chromosome carrying the Tripsacum centromere is shared between the two maize centromere-carrying chromosomes, approximately equally. At pachytene in type b, in 24 cells with a trivalent configuration where analysis was possible, 19 contained homologous synapsis of the distal region only, 3 contained homologous synapsis of the intercalary region only, and 2 contained homologous synapsis of both distal and intercalary regions (with change of pairing partner). If this is a representative sample, most trivalent formation depended upon synapsis of the distal region in type b. Nevertheless, the frequency of both homologous synapsis and crossing over in type b corresponded to expectation from availability for these events of an extent equal to the combined potential extent within both distal and intercalary regions.
In the absence of some unrecognized artifact or sampling error, it is difficult to escape the conclusion that both synapsis and crossing over depend upon total extent of available homology, although most of the occurrences of these events seem to have been shunted into the distal region in type b. Is commitment to both synapsis and crossing over established at prealignment in a manner partially dependent on intercalary chromosome regions, although actual synaptic and crossover events tend to be distally located?
Marjorie Maguire
Return to the MNL 57 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}