For some years now there has been some interest in leaf striping and its rela- tionship to mutator (Mu) activity (MGCNL 55:2-3, 1981; 56:2, 1982). As was mentioned in the 1981 report, considerable striping was observed in Mu stocks grown in Berkeley. These plants were scored on a six-step scale from 1 (no striping) to 6 (heavily striped), self-pollinated, and outcrossed to a standard line. The outcross progeny were selfed and scored for the segregation of new mutants and the mutation frequency calculated for each outcross. The seedling classifications were made without knowledge of the striping classifications. After the mutation frequencies were determined, the striping determination for each tested plant was obtained and the coefficient of correlation (CC) was calculated for the frequency of mutants and the degree of striping. Two sets of plants were
tested. One set was derived from striped mutator stocks that Dr. Michael Freeling had developed and the second consisted of some of my Mu stocks which had been previously grown in Iowa and had shown no striping. At Berkeley, striping was observed in some of these I.S.U. stocks.
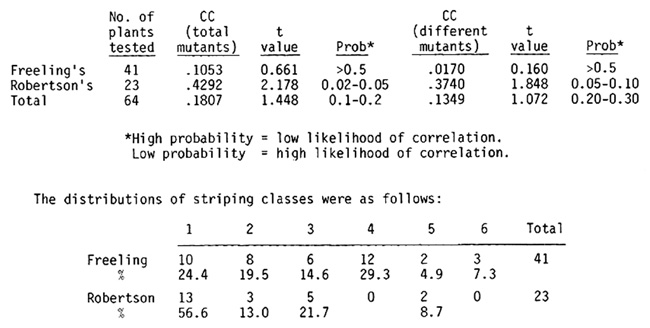
The correlation results are summarized below:
The distributions of striping classes were as follows:
In Robertson's material the percentage of nonstriped plants was twice that of Freeling's, and there was low correlation of striping with mutator activity. In Robertson's stocks, although fewer plants striped, there was a correlation of mutator activity with striping.
I believe the difference in these two stocks can be explained on the basis of what is known about their origin and environmental factors influencing striping. When Freeling grew the original Mu stock I sent him at Berkeley, he observed striping. Assuming the striping was the result of Mu activity, he selected these plants for crossing and continued to select for striped plants (= Mu plants?) in future generations. Thus he had selected strongly striped stocks. Striping was rare in Robertson's stocks grown in Iowa and Hawaii and no selection for the striping phenotype was made.
In last year's News Letter (MGCNL 56:2, 1982) it was reported that low temperature will cause striping in not only Mu stock but also normal lines. With these facts in mind, the following might explain the experimental results. The growing conditions (i.e., low temperatures) at Berkeley are such that in most years striping will be induced in many plants, normal or Mu. Perhaps Mu plants are more susceptible to striping at low temperature (this is yet to be established, but if true, it would not be inconsistent with a viral infection, since viral symptoms are sometimes more extreme at lower temperatures). Thus, Mu plants grown at Berkeley may have two types of stripes: Mu-induced and those induced by low temperature. If, over several generations, selection for striping was carried out at Berkeley for plants that were more prone to cold-temperature-induced striping, then there would be little correlation expected between striping and Mu. On the other hand, Robertson's stocks were never subject to selection for cold-temperature-induced striping. Thus, when grown at Berkeley, more of the striping might have been due to the Mu response to cold temperature relative to the normal cold-temperature-induced striping, and hence the stronger correlation with striping.
These results suggest that striping might be a phenotypic expression of Mu. However, one has to be aware of the low temperature effect when trying to select Mu plants on the basis of striping (e.g., among Freeling's material there were two striped plants that showed no mutator activity). Before striping can be used as a diagnostic guide for Mu it will be necessary to better characterize cold temperature striping as opposed to mutator striping, if indeed they are different phenomena. The fact that most Mu progeny are Mu will complicate the analysis because the chance of picking a Mu plant each generation, regardless of phenotype, is high. In my early work with mutator, this transmission pattern led me to wrongly conclude that mutator was associated with y9, since selection and propagation were made on the basis of y9 being present.
There is one additional observation that bears on the striping phenomenon. This last spring was unusually cool in Iowa. My first planting (April 30) germinated well, but after the two to three leaf stage there were a couple of weeks of cool weather. During this time little growth took place and there was extensive yellowing of the leaves. A few weeks later, for the first time in my field, I saw considerable striping in my Mu lines. Similar striping was not observed in my second planting (June 2). In checking with the plant extension workers on our campus, they informed me that they were deluged with calls from farmers this spring about striped plants in their fields. They further informed me that striping is a commonly observed phenomenon in cool springs, and that there is considerable varietal variation with regard to susceptibility to striping.
I have talked to our corn breeders here and they report seeing such cold temperature striping, but they are not aware of any studies on this phenomenon. If anyone knows of reports and/or studies on this effect of low temperatures, I would appreciate being made aware of them.
Donald S. Robertson
Return to the MNL 57 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}