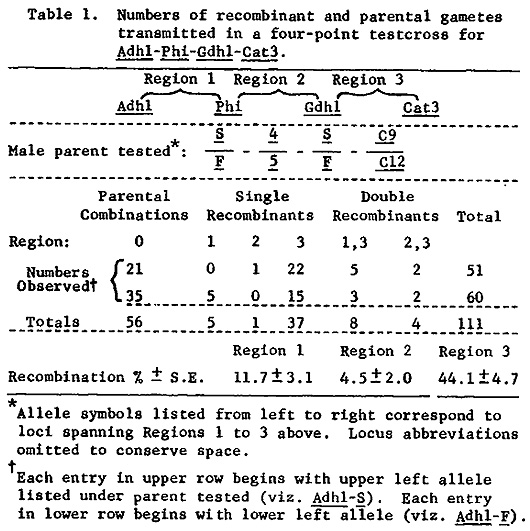
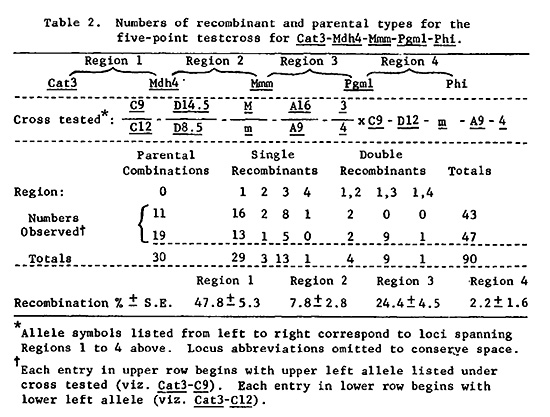
We recently reported recombination values for several multiple point test-crosses involving Gdh1, Phi, Adh1, Pgm1, and Mdh4, all on the long arm of chromosome 1 (Genetics 96:697-710). We also routinely score most of our materials for Cat3, reportedly localized to the same chromosome arm by Roupakias et al. (TAG 58:211-218). In the course of determining the amount of recombination between Cat3 and isozyme loci spanned by Gdh1 and Mdh4, we found that Cat3 was at most loosely linked to any of these loci. We used a genetic stock obtained from Ed Coe to make two four-point testcrosses for Cat3, Gdh1, Phi, and Adh1. The same male parent (2323-17) was used for crosses with 4Co82 and 1873-4 (the latter is one of our standard tester stocks derived from Co125 x H25). The male parent had genotype Cat3-9/12, Gdh1-F/S, Phi-4/5, and Adh1-F/S. The male gametes transmitted are listed in Table 1. To test for Cat3 linkage to Mdh4, we used a testcross of Phi-3/4, Pgm1-A9/A16, Mmm-M/m, Mdh4-D8.5/D14.5, Cat3-C9/C12 as male onto W629A. The male parent had been experimentally introgressed with teosinte (source: race Central Plateau, collection K69-2) and the segment of 1L on which Pgm1-A16 and Phi-3 resided was of teosinte origin, while the Mdh4-D14.5 to Mmm-M segment was of maize origin. Use of such a stock might be expected to decrease the amount of recombination between Mdh4 and Cat3 if there were any lack of chromosome homology between maize and teosinte in a region of 1L spanned by the two loci. The results are listed in Table 2. Clearly Cat3 is not closely linked to Mdh4. The recombination values for Regions 2 and 3 correspond closely to those estimated earlier (7.8 ± 2.8 vs. 4.4 ± 1.1 and 24.4 ± 4.5 vs. 19.0 ± 2.1). For Region 4, however, there is a striking discrepancy. Here we have only 2.2% recombination, while a similar cross involving standard inbred lines resulted in 19.5% recombination. Presumably this discrepancy is caused by lack of sequence homology between maize and this accession of teosinte, perhaps as a result of an inversion covering most of Region 4. The only evidence these data hold for Cat3 belonging to this linkage group is the lack of C9-D8.5-m-A16-3 type double-crossover gametes in Table 2. About 5 were expected, none were observed. Yet 9 of the complementary type double-crossover gametes (C12-D14.5-M-A9-4) were observed.
M. M. Goodman and C. W. Stuber
Return to the MNL 56 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}