Two sets of sublines of the inbred A344 were included in the experiment. The check set was composed from 10 sublines (CP1 - CP10), randomly selected from the check population. The experimental set included the check subline CP1 and 9 NMU induced macromutants (MP1 - MP9; 3 semi-dwarfs, 2 dwarfs, two-tassel, prolific, tall stalk, tassel seed-2 forms). The major phenotypic changes were controlled by recessive genes in all mutants. The lines of each set were diallel-intercrossed within their sets in one direction. The parental forms as well as hybrids were included in trials utilizing one-row plots, 4.9 m2, with 20 plants in the plot, with six replications. The combining ability for grain yield and plant height was analyzed according to Griffing (Method 4, Model 1). The results are the following:
1) Mutants were lower than the check sublines in grain yield (19.7 q/ha vs. 27.0 q/ha).
2) Hybrids including mutants also show a marked grain yield reduction. The mean grain yield of the control set was 34.7 q/ha, and that of the mutant hybrids was 29.8 q/ha. However, the minimal yields within each set were practically the same, 25.0 q/ha and 23.8 q/ha, respectively. The range of variability was reduced from 23.0 q/ha to 17.0 q/ha.
3) The reduction of the mean plant height of mutant hybrids was significant in comparison with that of the control set (130.2 cm vs. 138.4 cm).
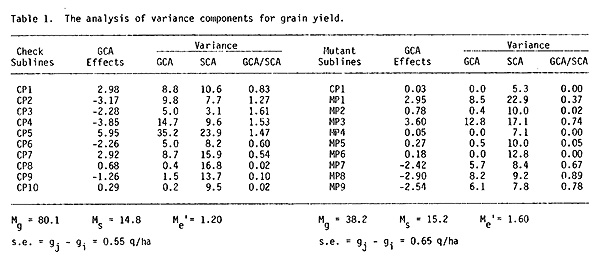
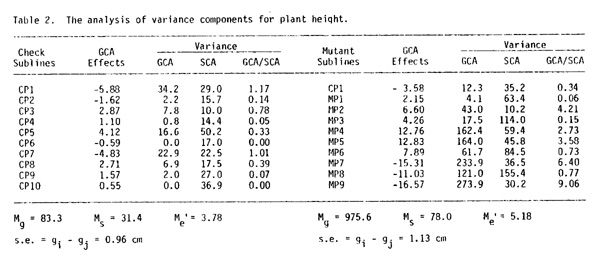
4) The grain yield and plant height variation in the set of hybrids obtained by crossing the control sublines were mainly determined by general combining ability (GCA). For the first trait the mean square deviation due to GCA (Mg = 80.1) more than by five times exceeded the mean square deviation value under the influence of the special combining ability (SCA) (Mg = 14.8) (Table 1), and for the second trait it exceeded more than two times (Table 2).
5) The grain yield reduction is explained by the reduction of the effects of favorable additive factors (Table 1). The GCA mean square value was reduced twofold in mutant hybrids comparing with the check set. The effect of specific factors was maintained at the same level.
6) In the control set the subline CP1 enters the best group for the GCA effect, and in the crosses with mutants it did not express its potential (Table 1).
7) Twofold increase in plant height variation range in the set of mutant hybrids is explained by a significant rise in effect of additively acting genetic factors.
8) The M3 plants both heterozygous and homozygous for dominant alleles of oligogenes did not differ in grain yield or stalk height. The hybrids from crossing CP1 and CP8 with a homozygous mutant and non-mutant plants from M3 families were also similar in these traits. It proves that oligogenes in a heterozygous state did not affect the vigor of A344 line plants.
From the published data we see that not all oligogenes in a heterozygous state express influence on the plant vigor. Many investigators consider that the differentiation of the inbreds is essentially associated with the accumulation of micro-mutations. Palenzona (MGCNL, 1968, 42:12-14) stated that an autodiploid inbred under the influence of spontaneous mutagenic factors showed a change of combining ability for plant height on account of additive genes.
We figure out two mechanisms leading to the essential change of additive genetic effect expression.
Hypothesis I. The polygenes acting additively differ from the polygenes acting non-additively in higher mutability.
Hypothesis II. Both polygene types display similar mutability. However, a maize genotype contains an insignificant number of loci that are able to interact specifically.
Let us divide the additive factors into two groups according to their resistance to mutagenic agents: (a) stable additive genes (SAG); and (b) mutable additive genes (MAG). This assumption is based on the widely accepted view about the "hot points." Let us assume further that the genes showing the controlling effect on the combining ability for grain yield are more stable than those controlling the plant height.
In this case all above mentioned peculiarities of the grain yield variability may satisfactorily be explained by an increase in concentration of recessive alleles of the MAG group in the mutant hybrid population. The frequency of undesired recessive alleles of minor genes was higher in mutants compared with check sublines. As a result of it the undesired effect of recessive genes is seldom compensated by the dominant genes of the partner in the cross. Respectively, the range of variability in plant height increased under the influence of new genes from the SAG group and new non-additive genes.
The genotypes of the check sublines utilized in this experiment have been developed under the influence of the following phenomena: (a) the long-term process of accumulation of spontaneous mutations, (b) the gene recombination process, (c) genetic drift. The valuable information does not allow a direct assessment of the factors governing the formation of specifically acting gene complexes. However, comparing the GCA and SCA variances in the check sublines and induced mutants we see that this process proceeded independently of the direction and intensity of variability of additive factors.
Thus, the analysis of the experimental data shows differences in quality and relative autonomy of additively and non-additively acting polygenes. In 1942 G. F. Sprague and L. A. Tatum found the phenomenon of relative independence of selection for GCA and SCA. It seems that the hypothesis of different nature of the genes controlling these traits may help in explanation of selection independence.
Acknowledgement: These experiments were realized due to the valuable help and supervision of my teacher, the late Academician M. I. Hadjinov.
A. Mashnenkov
Return to the MNL 56 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}