Since the first reports of Aberrant Ratio (AR) by Sprague and McKinney (Genetics 54:1287-1296, 1966; Genetics 67:533-542, 1971) there have been divergent opinions regarding the cause of this phenomenon. Although much of the data suggests that a gene regulatory mechanism is responsible, other bases have been proposed.
In the 1981 Newsletter, Nelson presented evidence on Aberrant Ratio involving the A and Su loci in stocks he obtained from Sprague. In two cases of the A a* type distortion (an excess of colorless kernels) in sib matings between plants from colored and colorless kernels, Nelson found that segregation of c in one case and c2 in the second accounted for the observed ratios. In a third instance which produced 61% colored kernels in sib crosses of plants presumably A a and a a, all other known aleurone color loci proved to be homozygous dominant. These plants occurred in a line which showed the normal 1:1 ratio of colored and colorless progeny in the previous generation. Nelson indicated that a gametophyte factor could account for these observations.
In another case involving distorted ratios of the Su su* type at the sugary locus, Nelson conducted self pollinations of presumaby Su su plants as well as outcrosses to Golden Cross Bantam. In those cases where an excess of sugary kernels occurred in the self progeny, an excess was also observed in the outcross although the excesses were greater than expected in the self than in the outcross populations.
In the following generation of one of these plants which gave 37% sugary in the self progeny, reciprocal crosses were made between plants of nonsugary kernels from the selfed ear and individuals from the Golden Cross Bantam line. In these crosses, ratios of approximately 1:1 were obtained when the nonsugary plants were used as females but excesses of sugary occurred when they were used as males. Nelson pointed out that a factor on chromosome 4 affecting pollen viability could account for these results.
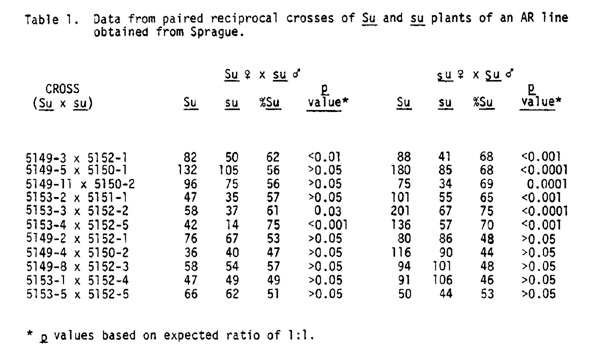
For the past two summers we have been examining the Aberrant Ratio phenomenon in some of the preexisting lines as well as in stocks generated from virus-infected plants in this laboratory. One stock obtained from Sprague (75:287-6 x 286-1) exhibited ratio distortions of sugary. In 1980, paired reciprocal crosses were made between sibling plants from Su and su kernels. Although seed set on these ears was low, the data were more than sufficient to draw some conclusions. The results of these pollinations are presented in Table 1. In six of the cases, there was an excess of the dominant phenotype in progeny of Su plants used both as males and females while in the remaining five, a 1:1 ratio occurred in all crosses. In five of the six instances where excesses of the Su phenotype were observed, the deviations were greater in progeny of the male flowers than in those of the female, while in one example the opposite was true.
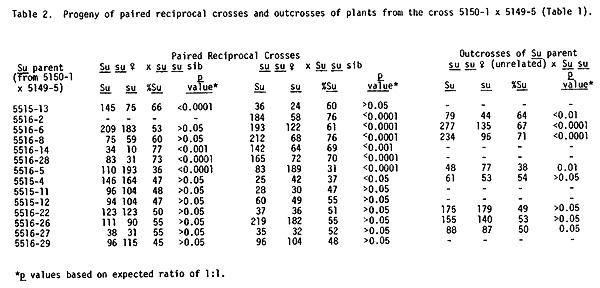
In the summer of 1981, further tests were conducted with progeny of the cross of 5150-1 x 5149-5 which generated 68% Su kernels. In these crosses paired reciprocal pollinations were made between plants from Su and su kernels of this stock and in addition, outcrosses of the Su individuals as males to an unrelated sugary tester were carried out. The results are listed in Table 2. In the first six cases, an excess of Su kernels occurred in progeny of both paired reciprocal crosses and in the outcross, where data were available. In seven of the cases, ratios were about 1:1 but in one instance (5516-5), an excess of the recessive phenotype was observed.
In most progenies which showed an excess of either the dominant or recessive phenotype, the deviations from a 1:1 ratio were greater when a plant was used as a male parent than when it was crossed as female. These kinds of transmission patterns are like those observed when a small chromosomal deletion has occurred. Rhoades & Dempsey in their studies of Inversion 3a (Amer. Jour. Bot. 40:405-424, 1953) found that some ovules possessing a chromosome 3 lacking the terminal 5% of 3L did function but that pollen grains carrying the same deficiency could not achieve fertilization in competition with normal grains. In studies of x-ray induced alterations at the A locus, Stadler and Roman (Genetics 33:273-303, 1948) found that deletions (a-X1, a-X2 and a-X3) were always transmitted at a higher frequency through ovules than through pollen. And, in a similar study involving markers on the short arm of chromosome 9, deficiencies including bz alone, both sh and bz, and one encompassing the c bz region all passed through the female gametophyte at a higher frequency than through the male parent (Mottinger, Genetics 64:259-271, 1970; and E. H. Coe, MNL 55:27, 1981).
The results of these studies along with the occurrence of ovule and pollen abortion caused by duplicate deficient chromosomes in translocation and inversion heterozygotes indicate that most chromosomal segments contain genes essential for normal gametophyte development. They also demonstrate that the pollen is affected to a greater extent by small chromosomal lesions than the embryo sac.
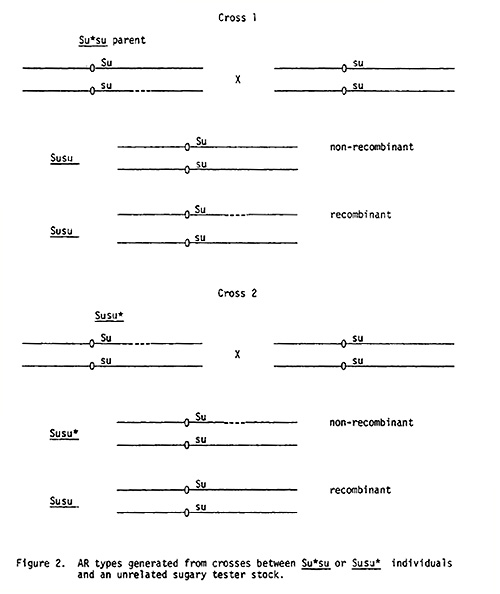
Assume for the sake of argument that in the stocks exhibiting ratio distortions of the sugary alleles, a small segment of chromosome 4 has been deleted or repressed. (Functionally, the results would be the same.) Then the results could be explained by the scheme diagrammed in Figure 1. According to this hypothesis, the Su* su type of distortion in the second generation would require that a normal chromosome 4 carrying the Su allele come from one parent while a chromosome 4 containing su and the affected segment be contributed by the other.
This can be tested since plants, presumably Su su, from lines exhibiting both the Su* su and Su su* type distortions have been crossed with an unrelated su tester which should not carry a deleted or repressed segment on chromosome 4. In the progeny of these heterozygotes backcrossed to the sugary stock, certain types of distortions should not occur. The chromosomal constitutions of these plants and their offspring are depicted in Figure 2. This theory predicts that distorted ratios of the Su* su type would not occur in the following generation of an Su* su or Su su* plant mated with a normal sugary tester. If they do, then other explanations, or modifications of this one, must be put forth.
Other observations from these crosses may also be possible. Since the AR stocks have been outcrossed to an unrelated line, the resulting hybrids should be considerably more vigorous than the former plants maintained by sib matings. If seed set is good in the backcrosses, then when the putative heterozygotes from the AR stock are used as females, ears showing off ratios should exhibit ovule abortion at frequencies which would account for the difference in the number of dominant and recessive kernels, if the deletion/repression model is correct.
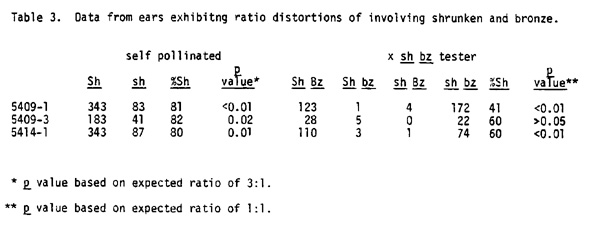
In addition to the AR stocks obtained from Sprague, ears exhibiting off ratios involving both sh and bz have been generated from virus infected plants in the summer of 1980 in Berkeley and in this laboratory. Plants of Mike Freeling's Adh1-S Adh2-P line, which are Sh Bz Wx pr A A2 c r in genotype, were infected with BSMV and crossed reciprocally to a stock of the genotype Adh1-F Adh2-N sh bz wx pr A A2 C R. The Adh1-S2P line is particularly susceptible to the virus and under the proper conditions plants with severe symptoms extending throughout all the leaves and which shed adequate amounts of pollen can be obtained. Over 2,000 F1 seeds from these crosses were planted in the summer of 1981 and the resulting plants were self pollinated and backcrossed as females to the sh bz wx tester. Ears from 174 of these plants have been analyzed to date and three exhibit ratios involving both sh and bz which deviate significantly from the expected values. The data from these three plants are presented in Table 3. Since the F2 progeny from the self pollinations segregate for colored and colorless kernels because of c and r, and since many of the colorless individuals cannot be distinguished from bronze seeds on these ears, only the Sh and sh phenotypes are recorded. On the backcross ears where colorless is not expressed, data on phenotypes of both shrunken and bronze alleles are listed. In all three cases, a significant excess of kernels expressing the dominant phenotype occurs on the F2 ears while in the backcross progeny, different results are obtained. In one example (5409-1) there is a significant deficiency of Sh seeds while in the second (5409-3) there is an excess, but the population is too small to derive any meaningful data. In the third instance (5414-1), there is a significant excess of Sh kernels. On the first and third backcross ears where populations are adequate, there is also a deficiency or excess, respectively, of Bz kernels.
To account for the data on off ratios of shrunken and bronze alleles by hypothesizing a mechanism in which gametophyte factors reduce the transmission of a particular homolog, a model must be presented which is more complex than the one used to explain the distorted ratios of sugary alleles. Since the sh and bz phenotypes are deficient on the three self-pollinated ears and in one of the backcross progeny, gametophyte factors could have been segregating in the sh bz tester stock. However, since so few plants were observed showing off ratios, the factors would have to be expressed only infrequently. And, to accommodate the deviations in opposite directions of the self and backcross ears of plant #5409-1 a means of transferring these factors from one homolog to the other somatically (as suggested by Brink et al., Science 159:161-170, 1968, to explain paramutation) must be invoked. The same kind of element could have arisen in the virus-treated parent and in the F2 plant, been transferred to the opposite homolog.
In addition to questions arising from these data, others must be answered as well. For example, in the studies reported by Nelson in which the c and c2 alleles were found to be segregating as well as a in AR lines obtained from Dr. Sprague, where did they come from if the original parents were a A2 C C2 R and A A2 C C2 R?
All of the data collected so far in various laboratories can be explained by a mechanism or mechanisms involving gene regulation; but clearly, a great deal more data are needed before the controversy surrounding the phenomenon called Aberrant Ratio can be resolved.
We thank Dr. Sprague for kindly providing stocks exhibiting Aberrant Ratio.
John Mottinger, Patrick Keller and Lisa Campopiano
Return to the MNL 56 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}