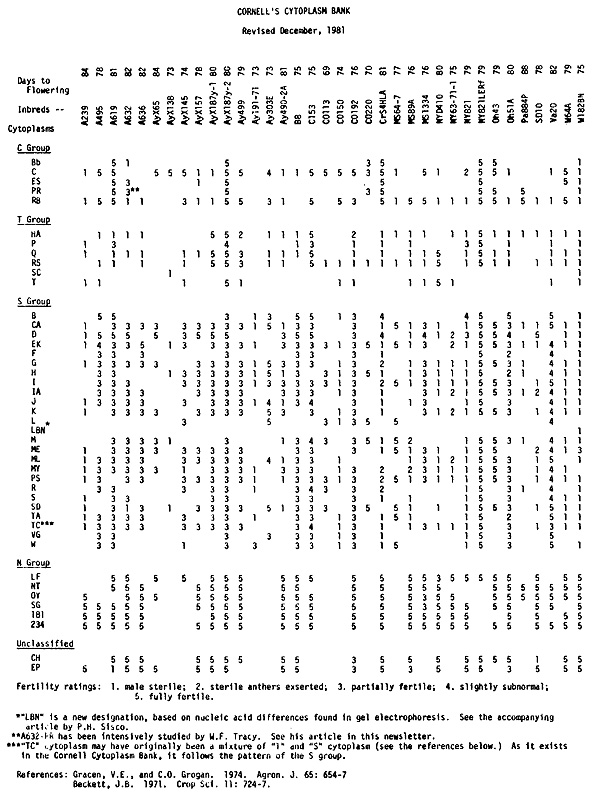
The accompanying chart lists the most recent visual ratings for fertility restoration for the 36 inbreds in 44 cytoplasms which are included in Cornell's Cytoplasm Bank. These numbers are based on ratings given the last two times these plants were grown out, namely 1975, when the entire set was rated by C. E. Manchester, and 1980 and 1981, in each of which half the set was rated by V. E. Gracen. The origin of the bank is described in the two references at the bottom of the chart (Gracen and Grogan, 1974, and Beckett, 1971). Each inbred/cytoplasm combination represents a minimum of five backcrosses and in a few cases as many as 14 backcrosses to the inbred, the average being 8 to 9.
To properly interpret the ratings, several important facts must be kept in mind:
1) Although every effort has been made to plant, harvest, shell, and package this material carefully, mistakes are possible. Given the thousands of opportunities for error, we feel that rather few have in fact been made and that these are usually easily traced. Nevertheless, anyone undertaking critical work with this material should check its authenticity with as many as possible of the techniques now available: (a) restoration patterns (available in the chart); (b) susceptibility to H. maydis, Race T or its toxin, a characteristic of members of the T group; (c) presence of the S-bands, which seem completely correlated with members of the S group (see Kemble and Bedbrook, Maydica 24:175, 1979, or Kemble, TAG 57:97, 1980); (d) presence or absence of other low-molecular-weight nucleic acid species (Kemble and Bedbrook, Nature 284:565, 1980); (e) patterns of mitochondrial and/or chloroplast DNA digested with restriction endonucleases (Pring and Levings, Genetics 89:121, 1978); (f) in vitro translation products of mitochondrial DNA (Forde et al., Genetics 95:443, 1980). In a few cases, the restoration data and the biochemical tests are not well correlated (see the problem of B, D, and ME cytoplasms in the accompanying article on diversity for restoration within the S group).
2) Even though enough backcrosses have been made to each inbred to make the plant types very uniform within a single inbred, nuclear heterogeneity persists. This heterogeneity has at least two causes: (a) linkage slows the return to homozygosity in a backcrossing program, even when the recurrent parent is completely homozygous; (b) no corn inbred is likely to be homozygous for all genes. Thus, even the recurrent parent is diverse in nuclear background (see the accompanying article about A632-Pr by W. F. Tracy). The result of this nuclear heterogeneity is that some of the diversity in restoration patterns within a group (C, T, or S) may be due to nuclear rather than to cytoplasmic differences. Residual nuclear diversity is the most likely cause of the bifurcated plants which occasionally appear in EK cytoplasm in Cornell's bank. The original source of EK cytoplasm was a male-sterile bifurcated plant in the variety "Early King" (A. L. Hooker, personal communication). The bifurcation has been carried along as an occasional adjunct to the sterility. Preliminary analysis of the bifurcation shows it to have a nuclear rather than a strictly maternal inheritance pattern.
3) Environment has a strong influence on the degree of male sterility and thus on the fertility ratings. A cytoplasm/inbred combination may be rated a "1" in one location or year and a "3" in another. S-type male steriles are especially variable. Our chart reflects ratings made in one location (Aurora, New York) during two seasons. In cases where an inbred-cytoplasm combination showed variability for restoration, the maximum rating is given.
P. H. Sisco, V. E. Gracen, C. E. Manchester and H. L. Everett
Return to the MNL 56 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}