The nondisjunction of B-A chromosomes provides a useful method for varying the dosage of specific chromosome segments of corn. However, the method is only applicable to A chromosome regions that are distal to B-A translocation breakpoints. Proximal regions cannot be duplicated by nondisjunction. Nevertheless, it is possible to duplicate proximal regions using B-A translocations. The procedure requires the production of double translocation chromosomes carrying one B-A interchange in the long arm and one in the short arm.
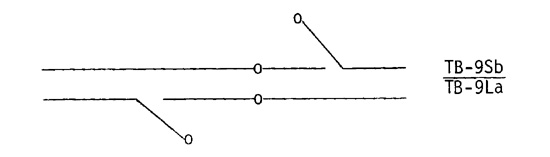
A double translocation was derived for chromosome 9 using TB-9Sb and TB-9La. The two translocations were combined in a single plant, producing a double heterozygote with one 9-B from each translocation and one B-9 from each. Complete pairing in meiosis is as follows:
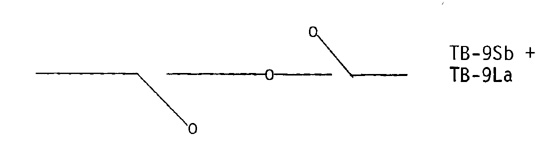
A double translocation was extracted from crossing over between the two 9-B chromosomes:
The proper combination of crossing over plus meiotic disjunction required to produce the double translocation occurs infrequently. Therefore, a selective method for identifying it was necessary. The simplest method of selection might be identification of double nondisjunction using 9S and 9L markers. However, in this case a modified TB-9Sb was used that eliminated the need for 9L markers (Translocation 2150; see Ch. 44 in Maize Breeding and Genetics, 1978). The translocation lacks the distal tip of the 9-B chromosome, which contains a factor required for nondisjunction. In the double heterozygote (TB-9Sb-2150/TB-9La), crossing over can produce a double translocation which contains the missing nondisjunctional factor. Only the double translocation is capable of giving nondisjunction of the B-9 from TB-9Sb. The double heterozygote was crossed as male to a bz bz tester and kernels with the bz phenotype were selected. These contained the double translocation. Some selection in progeny crosses was required to make certain that the B-9 from TB-9La had not been lost by nondisjunction.
Once the double translocation was established, it was crossed as male to a bz bz wx wx B B Pl Pl tester to select double hypoploid plants lacking both B-9's. Progeny with the bz plant phenotype were selected, since they lacked the B-9 from TB-9Sb. Among these plants, root tip chromosome counts were used to select plants with the 9 9-Ba+b constitution. Meiotic samples from 9 9-Ba+b plants frequently showed a lack of pairing between the chromosomes at diplotene. This is not surprising considering the evidence of Burnham et al. (Gen. 71:111-126, 1972) that corn chromosomes initiate pairing near their distal ends. The 9 and 9-Ba+b have no ends in common. The frequent lack of pairing in the double hypoploid allowed recovery of the 9-Ba+b chromosome in crosses of the type 9(wx) 9-Ba+b(Wx) X wx wx. Migration of the univalent 9 and 9-Ba+b to the same pole resulted in viable Wx progeny (9 9 9-Ba+b). By itself, the 9-Ba+b is lethal due to deficiencies.
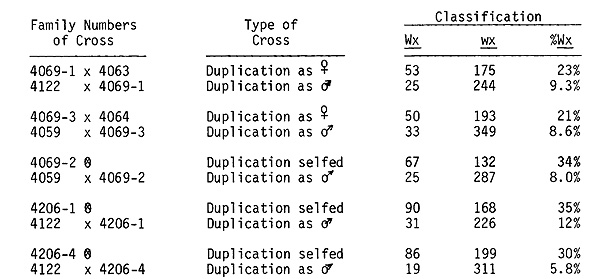
In 9 9 9-Ba+b plants, the central region of chromosome 9 has been duplicated. This duplication may be useful in experiments which involve gene dosage. Recent tests of transmission of 9-Ba+b indicate that a homozygous duplication of the chromosome can also be produced. The Wx allele is a precise marker of the 9-Ba+b chromosome since it virtually never recombines with the normal 9. It is located at the distal tip of the 9-Ba+b short arm. Several 9 (wx) 9(wx) 9-Ba+b(Wx) plants were crossed as male and female to a wx wx tester. Other such plants were crossed as male to a wx wx tester and self pollinated. Transmission of Wx in these crosses is given in the following chart:
The extra (Wx) chromosome is transmitted at a reasonable rate through both male and female parents. It should, therefore, be possible to select 9 9 9-Ba+b 9-Ba+b plants from the self pollinations. Selection can be made by identifying plants with all Wx pollen. It is also possible that the homozygous duplications will be stable and breed true. This would be expected if preferential pairing between structurally similar chromosomes is 100% and bivalents are always formed. Whether or not a stable 22-chromosome line can be produced depends on the mechanism of chromosome pairing. A stable line should be possible if chromosome pairing: (a) always begins in distal chromosome regions, and (b) synapsis of internal regions always follows, and (c) independent pairing of proximal regions always occurs last. Once a homozygous duplication line is established, its stability will be tested using Wx transmission tests and cytological examinations of bivalent pairing at diplotene.
W. R. Carlson
Return to the MNL 56 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}