Chromosome aberrations in autotetraploids create much more complex situations than they do in diploids. The relative rates of formation of various types of aberrations, the pairing configurations, the patterns of chromosome disjunction, and the fates of deficient chromosomes and unbalanced spores and zygotes are all different on the tetraploid level.
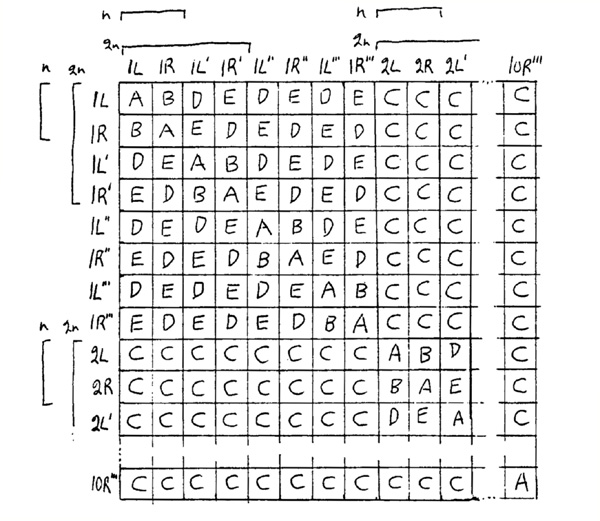
First, the difference between the relative rates of different aberrations may be explained by the table below, which gives the results of 2 break events. One break's position is given on the horizontal row and the other break's position is given on the vertical column. The symbols L and R refer to the left and right arms of the chromosome. To distinguish between homologous chromosomes, primes are used. Thus, 1L, 1L', 1L", and 1L"' are four homologous left arms of chromosome 1. This model uses a species with n=10, so there are 80 arms where breaks can occur and 6,400 possible combinations (3,160 are duplicates). A diploid would have 40 entries on each side and 1,600 combinations. A haploid would have only 20 entries and 400 combinations.
This table assumes that breaks occur at random, that broken ends unite at random, and that each arm has the same rate of breakage. Because unions take place only between broken ends that are very close together, non-random positions of chromosomes will affect the relative frequencies. If homologous chromosomes tend to be associated then there will be a greater frequency of D and E events. Also, the longer arms will have more breaks than short arms. However, the table gives an approximation.
The relative frequency of reciprocal translocations is not affected by ploidy, while the frequencies of the inversions decrease and tandem duplications and pseudo-isochromosome production increases with higher levels of ploidy.
The following discussion is only a brief summary of the effects of various aberrations in autotetraploids.
Deficiencies. Most deficiencies cause lethality in haploid gametophytes and cannot be maintained in diploids. However, in an autotetraploid they can be transmitted through the gametophyte generation if they are accompanied by a complete chromosome. They persist for a time in autotetraploid populations. If there is a random mating population with an initial frequency of a deficiency (Df) of .25, after 10 generations it will decline to .0426. This assumes random chromosome pairing and an alpha value of 1/7 (equivalent to random chromatid assortment). Preferential pairing may occur in NNDfDf plants. An important pairing site may have been deleted from the Df chromosome. Preferential pairing would reduce the rate of Df elimination from the population.
Dicentrics. This type of aberration is produced very frequently (C, D, and E events). However, it is very rarely found. It is believed by Steinitz-Sears and Sears (Genetics 42:623-630, 1957) that they generally break and give rise to two chromosomes with terminal deficiencies.
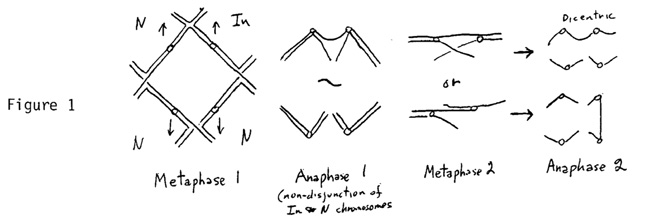
Paracentric inversions. Crossing over between normal and inversion chromosomes in the inverted region causes the formation of a bridge which undergoes a bridge-breakage-fusion cycle to form deficient or duplicate-deficient chromosomes. On the female side in diploids this bridge tends to orient chromatids so that only a complete inversion or normal chromatid goes to the functional megaspore. However, in an autotetraploid this bridge may not be resolved at the first or second division of meiosis, as shown in Figure 1.
The deficient or duplicate-deficient chromosomes are eliminated from the population. If the initial frequency of an inversion is less than 50% in a random mating population it will be eliminated. This is a result of selection against the heterozygote which fixes the gene (and chromosome arrangement) which was in the majority. The rate of elimination will depend on the frequency of crossing over in the inverted region and the amount of preferential pairing (which affects the amount of pairing between the inverted and normal segments and affects chromosome disjunction patterns). There are 12 different genotypes to be considered: NNNN, NNNIn, NNInIn, NInInIn, InInInIn, NNNDf, NNDfDf, InInInDf, InInDfDf, NNInDf, NInInDf, and NInDfDf. The Df chromosomes are of different types; some resemble the inversion chromosome and some the normal chromosome. Consequently, the expected gametic outputs of genotypes with Df chromosomes are difficult to predict because of different amounts of preferential pairing.
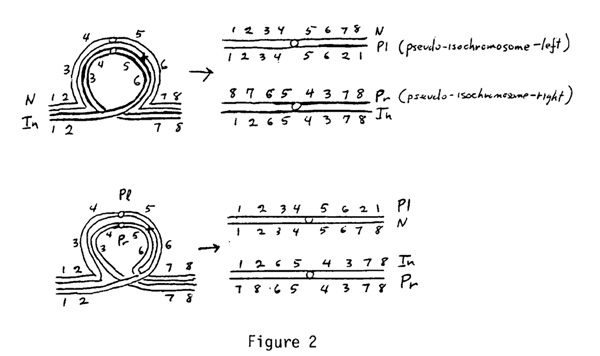
Pericentric inversions and pseudo-isochromosomes. Crossing over in the inversion chromosomes produces two duplicate-deficient chromosomes that are the same as pseudo-isochromosomes that are produced by radiation where opposite arms of homologous chromosomes have exchanged segments. In a diploid population a pair of pseudo-isochromosomes do not persist. They pair at meiosis and they separate. They cannot survive without each other or a complete chromosome. In an autotetraploid they can persist for a time because they may be directed to the same pole at meiosis and enter the same gamete. The elimination of a pericentric inversion from a random mating autotetraploid population (where the pericentric inversion is in the minority) is confounded by the fact that if a pair of pseudo-isochromosomes cross over in the inverted region they will restore a normal and an inversion chromosome (see Figure 2).
Eventually Pl and Pr chromosomes will be eliminated as the lethal PlPl and PrPr gametes. As with the paracentric inversion we have a bewildering assembly of 27 genotypes (often with peculiar preferential pairing) to contend with: NNNN, NNNIn, NNInIn, NInInIn, InInInIn, NNNPl, NNPlPl, NNNPr, NNPrPr, InInInPl, InInPlPl, InInInPr, InInPrPr, NNInPl, NNInPr, NInInPl, NInInPr, NNPlPr, InInPlPr, NInPlPr, NPlPlPr, InPlPlPr, NPlPrPr, InPlPrPr, NInPlPl, NInPrPr, and PlPlPrPr.
Reciprocal translocations in an autotetraploid lead to extremely complex situations. If the disjunction of chromosomes from multivalents was all alternate, then a translocation could be treated like a gene from a population genetics point of view. In maize it is about half alternate and half adjacent disjunction. Adjacent disjunction in a diploid produces unbalanced spores that abort. In an autotetraploid many types of unbalanced gametes are viable.
For the sake of brevity and clarity the genotypes of gametes and zygotes will be symbolized as sequences of 4 numbers. Thus, if the translocation was between chromosomes 1 and 2, the first number would indicate the number of chromosomes 1 and the next three numbers would indicate the numbers of 2, 12, and 21 chromosomes, respectively. There are 19 possible genotypes for the gametes: 1111, 2200, 0022 (balanced), 2101, 1012, 1210, 0121, 2110, 1021, 1201, 0112, 2011, 1102, 1120, and 0211 (unbalanced) and 2002, 0220, 2020, and 0202 (lethal). The non-lethal gametes would combine to form 65 zygotic genotypes. Only 5 are balanced: 4400, 0044, 2222, 3311, and 1133. Sixty other genotypes (the reader will be spared a list of them) have various degrees of imbalance.
Assuming the ends of the chromosomes paired at random the pairing configurations of NNNT (3311) plants would be 1/9 2II+IV, 4/9 II+VI, and 4/9 VIII and that of NNTT (2222) plants would be 1/81 4II, 8/81 2II+IV, 15/81 2IV, 16/81 II+VI, and 41/81 VIII. While the gametic output of quadrivalents could be computed that of sexavalents and octavalents is more difficult as the patterns of disjunction may be irregular. There would probably be frequent 5-3 separations of the chromosomes of an octavalent. Even in diploids translocation quadrivalents split 3:1 frequently. Thus we would have to consider aneuploid gametes as well. Even just considering the pairing configurations and gametic output of 65 different types of numerically euploid genotypes is a stupendous task. Also it is difficult to hypothesize the effect of competition between balanced and unbalanced gametophytes and the differential viability and reproductive success of the balanced and unbalanced sporophytes. Preferential pairing is another factor to consider.
Thus any complete theoretical treatment of the population genetics of reciprocal translocations in autotetraploids where there is adjacent disjunction is the work of a fool or a charlatan.
Tandem duplications in diploids have a low transmission rate through the pollen if they are large enough to cause imbalance affecting their competitive ability. Consequently these duplications do not become established in diploid populations. However, in an autotetraploid there is a lot of aneuploid pollen, the imbalance is reduced, and thus competition may be less intense. Also, tandem duplications can be transmitted in combination with the internal deficiency formed by the same event.
The theoretical effects of chromosome aberrations in allotetraploids also have been studied. There are many similarities and the behavior of the aberrations in allotetraploids would shed some light on their behavior in autotetraploids. Also, another approach is to obtain parthenogenetic diploids from autotetraploids that are heterozygous for aberrations. For example, it should be possible to get PlPr plants and determine the rate of reversion to normal and pericentric inversion chromosomes.
G. G. Doyle
Return to the MNL 56 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}