In recent years we have been occupied with the study of the mechanism responsible for the loss of segments of chromatin from knobbed A chromosomes at the 2nd microspore mitosis. We were able to show that loss resulted from the rupturing or breaking of dicentric bridges. These dicentric bridges were the consequence of a delayed replication of the heterochromatic knobs. Any knobbed A chromosome was subject to loss. We also demonstrated that faulty replication of knobs only happened in microspores with two or more B chromosomes. An unexpected bonus or spin-off from these investigations on chromatin loss, in which bridge breakage played a cardinal role, was the appearance of three mutable systems. We did not anticipate their occurrence and our experiments were not designed to reveal their presence but crop up they did, somewhat to our annoyance, since they diverted our full attention from the main thrust of our studies. However, we could not completely disregard them and a considerable body of information has been obtained in recent years. This report is a summary statement of our progress to date in analyzing these mutable systems, all of which proved to involve a controlling and a responding element.
Origin: As stated previously, three new mutable systems were discovered in stocks undergoing the high-loss phenomenon. The first of these involved an a-m responding allele (tentatively designated a-m-rh to distinguish it from the a-m responders of McClintock, Neuffer and Peterson) and a mutator designated Mrh. Mutation from A to a-m occurred in an F1 (30418) from a cross of the high-loss stock with a high-knob strain. The F1 individual had 4 B chromosomes and numerous knobs (including K10) so it was well endowed with heterochromatin. It was homozygous for the A allele. The mutation was detected when the F1 was used as male parent in a cross to an a tester. All progeny kernels were colored as expected save an exceptional kernel which had a white endosperm with colored dots. The embryo proved to be a/a-m-rh; it was heterozygous for the new a-m allele. The mutator Mrh has been found in plants with the high-loss background, so its precise time and place of origin are unknown.
A similar situation was found with the second system, which involves a mutable bz allele in 9S. The mutator, Mut, was carried in the high-loss stock but its presence was not recognized until a mutation arose at the Bz locus to give a bz allele which was mutable with Mut. It is designated as bz-m-rh. This mutation occurred in a derivative of the same F1 plant described above. Following five generations of inbreeding and one outcross to an a tester, a plant arose which proved to be heterozygous for the new responding allele. Subsequent tests of one of the intermediate generations revealed a plant containing Mut that continues to produce Bz Æ bz mutations, presumably to mutable alleles. Since the Bz/Bz parent was crossed to a bz-m mut tester, it has not yet been possible to determine whether the recurring bz mutations are to stable or mutable alleles.
The third system involves the responder, bz2-m in chromosome 1, originating in Neuffer's stocks and obtained from the Maize Coop. We found a new mutator, called Ac2, which interacts with bz2-m, as does Ac in Neuffer's stocks. Ac2 was detected in a cross of the 30418 F1 male parent with the Coop. bz2-m stock (a stable bronze without mutators). The source of the Ac2 is not yet clear; the few plants of high-loss background that we tested did not contain Ac2 but it may have been present in the high-knob stock.
The fact that all three of the mutable systems were associated, either in the occurrence of the mutator or the inception of new responders, with strains containing a large amount of heterochromatin may be of some significance. The 30418 F1 carried 4 B's as well as numerous knobs, including knobs on chromosomes 3, 4, and 1. Crosses of the 30418 F1 as male to female parents with recessive genes on those chromosomes showed that the high-loss phenomenon producing chromosome breaks was occurring in the 30418 male parent. Instability in the genome caused by repeated cycles of breakage initiated at the second microspore division may have been responsible for the origin of the new mutable systems. Such an origin was postulated by McClintock and supported by her studies and those by Doerschug. They found new Dt-like mutations arose in a dt plant which had a chromosome 9 undergoing the breakage-fusion-bridge cycle.
Phenotypes and dosage effects: The unstable phenotypes detected in the high-loss pedigrees show some similarities: in all three cases, reverse mutations in the presence of the controlling gene occur late in endosperm development giving fairly uniform small dots of full color on a colorless or bronze background. With increasing doses of the mutator, dots are more frequent in number but the time of mutation is not affected. In the a-m, Mrh system, kernels with three doses of responder and three doses of mutator are almost completely colored since the numerous dots are confluent, while in the bz2-m, Ac2 system such kernels have an average of more than 300 dots per kernel. The latter system has been most thoroughly analyzed; counts of dots per kernel gave the following results:
The action in aleurone cells of Ac2 on bz2-m is strikingly different from the interaction of McClintock's Ac with the same responding allele. Ac2 in a single dose usually gives no Bz dots while her Ac induces early reversions to Bz resulting in a coarse pattern when one dose is present. With two doses of Ac, there are many late mutations and with three doses, mutations are few and very late (Neuffer, MGCNL 28 & 29). The virtual absence of Bz dots in kernels with one dose of Ac2 was at first considered to be due to inactivation of Ac2 in the male gametes. We no longer subscribe to this view. It is apparent from the huge difference in mutation frequency between Ac2/Ac2/ac2 and Ac2/Ac2/Ac2 endosperms that the Ac2 contributed by the pollen parent is not inactive. Male gametes with two doses of Ac2 arise by transposition and kernels with two paternally derived Ac2 alleles have a low rate of mutation. The data presented above could be interpreted as indicating a lessening of Ac2 activity when Ac2 is introduced through the male. However, modifying genes affecting mutation rate are known to exist. The reported difference between kernels with two maternally contributed Ac2's and those with two Ac2 alleles of paternal origin may only reflect the effect of different constellations of modifiers. In support of this possibility is the average dot number of 12 found in kernels of another strain which possess two paternal Ac2's. There is no compelling evidence of pollen inactivation of Ac2. One dose of Ac2 simply does not produce the threshold necessary to induce mutability of the bz2-m allele and the exponential increase found in going from two to three doses represents a synergistic dosage relationship.
The A, Bz and Bz2 loci all affect mature plant and aleurone pigmentation. Plants carrying the a-m Mrh and bz2-m Ac2 systems were examined for sectors of A or Bz2 tissue in the stalk, husks or anthers. No Bz2 or A sectors were found and it was concluded that mutations are not induced by Mrh or Ac2 throughout the life cycle but appear to be restricted to the endosperm and germinal tissue. In bz-m Mut plants, very fine Bz stripes were found in the anther wall of plants carrying appropriate plant color factors, but no stripes were observed on the stalk or husks.
Linkage of mutator genes: If Mrh undergoes transposition to a heterologous chromosome or to a new site in the same chromosome, obviously it cannot be assigned a permanent position in the maize genome. That transposition takes place is indicated by the finding of kernels with sectors differing markedly in number of mutations to A, but we have not yet determined its frequency. However, transposition is low enough that no difficulty is encountered in obtaining an accurate map position in a specific chromosome. Immediately following the detection of Mrh we placed it in the long arm of chromosome 9. Since Mrh can only be scored in kernels homozygous for a-m or heterozygous for a-m and a, the most informative matings are those testcrosses where all of the resulting kernels possess the a-m allele. Data from crosses of a-m/a-m or a-m/a; Sh Wx Mrh/sh wx mrh females by a-m/a-m;sh wx mrh/sh wx mrh males are summarized below:
The waxy translocation stocks derived by Anderson
and Longley were used in placing Mut and Ac2. Mut shows 32% recombination
with wx in the backcross involving T2-9b (breakpoint 2S.18) and no linkage
with waxy in backcrosses involving the other translocation stocks. More
precise location of Mut on chromosome 2 awaits results of our crosses with
chromosome 2 tester stocks. Ac2 crosses routinely give a low but variable
percentage of kernels with no dots even when the mutator is introduced
through the female parent. This complication must be kept in mind in the
analysis of linkage data. Nevertheless, linkage of Ac2 on chromosome 8
became evident in backcrosses with T8-9(6673) (breakpoint 8L.35), where
a recombination value of 19.8% was found for Ac2-Wx in one test involving
526 bronze kernels. In a second test, an apparent recombination of 23.3%
in a population of 304 bronze kernels was reduced to 12.5% when only the
positive mutable class was analyzed. Although the recombination frequencies
were variable, the data indicate linkage of Ac2 on chromosome 8. To further
locate Ac2, tests were made with the ms8 gene in chromosome 8. Plants homozygous
for bz2-m and heterozygous for Ms Ac2/ms ac2 were obtained. Self-pollinations
of these gave a 1:1 ratio for bronze mutable and bronze stable kernels
(653:669). The bz mutable kernels are either Ac2/Ac2/Ac2 or Ac2/Ac2/ac2
in constitution while the bz stable kernels are ac2/ac2/Ac2 or ac2/ac2/ac2
. As noted earlier, one dose of Ac2 does not elicit response from bz2-m.
Both classes of seed were planted and the ensuing sporophytes scored for
male sterility to give the following data:
| mutable bronze | stable bronze |
| 327 Ms 0 ms | 170 Ms 152 ms |
The ratio of Ms:ms is close to the expected 3:1. If Ac2 and Ms8 are completely linked, all plants from bz mutable kernels would be male fertile while a 1 Ms:1 ms ratio is expected from the bronze stable class of kernels. This proved to be the case. More significant is the fact that no male sterile plants came from bronze mutable kernels. The conclusion is inescapable that Ac2 and ms8 are very closely linked. The tight linkage of ms8 and Ac2 will be useful in determining the location of newly transposed Activators and in differentiating between the separation by crossing over of two tightly linked Ac's and a change in state. Tests with TB-8a have shown that ms8 is distal to the breakpoint at .7 in 8L. Our data with the T8-9(6673) show Ac2 is approximately 20 units from wx. If wx accurately marks the breakpoint at .35 in 8L, the consensus from all available data is that Ac2 lies in the distal .3 of the long arm of chromosome 8.
Interaction of mutator systems: The chart below summarizes the interaction of several maize mutators belonging to two-unit systems with a number of responding alleles. It is based partly on data presented in a similar chart (see Peterson, Maize Breeding & Genetics, 1978) and partly on tests performed in this laboratory. The + sign indicates a positive interaction and the - sign no response.
In most cases, there is a high degree of specificity in these systems, i.e., a particular responder at a given locus is activated by only one mutator and a particular mutator affects only one responder. The exceptions identified in the chart are in the horizontal row opposite Ac=Mp and in the vertical column under bz2-m. Ac interacts with Ds and with the bz2-m allele, while bz2-m is induced to mutate by both Ac and Ac2. Without information supplied by Ac2, it might be concluded that a standard Ds element is present at the bz2-m allele--i.e., bz2-m = Bz2 Ds. The differential interaction of the two responders (Ds and bz2-m) with Ac2 (+ with bz2-m and - with Ds) indicates that the suppressive element at the Bz2 locus is not identical to a standard Ds. Furthermore, the mutators Ac and Ac2 differ in their specificities. Unlike Ac, Ac2 has no effect on standard Ds, as indicated in the above chart, or in the wx-m1 (Wx Ds) kernels obtained from the cross of wx/wx-m1; Bz2/bz2-m; Ac2/ac2 female parents by wx/wx; bz2-m/bz2-m; Ac2/Ac2 males. Both the bz2 stable kernels (one dose of Ac2) and the bz2 mutable (three doses of Ac2) had stable waxy endosperms. Ac2 in one or three doses obviously did not interact with the Ds element associated with wx-m1 to give wx Æ Wx reversions.
Tests with wx-m9 (Wx Ac) revealed that no change in endosperm phenotype occurred in wx/wx/wx-m9 kernels having four doses of Ac2 (obtained by using a transposed Ac2). In summary, Ac2 has no effect on Ds in its standard position in chromosome 9 between Sh and Wx, fails to interact with the Ds element in wx-m1 and likewise does not affect the Ac gene in the autonomous wx-m9 (Wx Ac) mutable system. It will be instructive to see if McClintock's Ac controls autonomous mutability induced by transposition of Ac2 if indeed such occurs. Our present evidence leads us to conclude that Ac2 is not a modification or change in state of the Ac studied by McClintock but represents an independent controlling element which shares with Ac only the ability to induce mutations of bz2-m. Additional evidence supporting this conclusion is given below in summary statements.
1. One dose of Ac elicits interaction from a responding gene while one dose of Ac2 is ineffective.
2. A dosage effect on time of mutation is present with Ac but does not occur with Ac2.
3. Ac is active in mature plant tissues; Ac2 is not.
Transposition: Our observation that bz2-m kernels with one Ac2 have no dots, that those with two doses of Ac2 have a low number while those with three doses have a high mutation rate made it possible to screen readily for transpositions of Ac2 by looking for exceptional kernels with a mutation rate much higher than that in sibling kernels. For example, in the cross of bz2-m/bz2-m Ac2/Ac2 females by bz2-m/bz2-m ac2/ac2 males, the genotypic constitution of the endosperm of F1 kernels would be bz2-m/bz2-m/bz2-m Ac2/Ac2/ac2. They will have a low number of dots per kernel. The embryos of the kernels with low dot number will be bz2-m/bz2-m Ac2/ac2 and in testcrosses they produce ears with a ratio of 1 bz mutable (low dots):1 bz stable. Occasionally, however, kernels are found with a much higher number of dots. Two or three such exceptional kernels may occur on a single ear. If they are clustered together on the ear, it is assumed that they all arose from one premeiotic mutational event. One seed from a cluster and all solitary exceptional kernels were grown and testcrossed as the female parent to determine their genotypic constitution for Ac2. A ratio of 1 bz mutable (high dots):1 bz stable in a testcross of plants grown from these exceptional kernels indicates heterozygosity for two closely linked Ac2 genes while ears with 3 bz mutable:1 bz stable ratios are expected if two independent Ac2 genes are heterozygous. In the latter case, one-third of the mutable class should have a high dot number, since the kernels would possess four Ac's and two-thirds should have a low dot number, comparable to that of the mutable class in testcrosses of sibling plants with one Ac2 since both would have two doses of Ac2 in the endosperm. The bronze stable class would have no Ac2. Good agreement was found between observed and expected frequencies. If, following transposition, two Ac2 alleles lie adjacent to one another in the same chromosome and there is no or little crossing over between them, a testcross ratio of 1 high dotted:1 no dots is expected. Deviation from a 1:1 ratio of high dots:no dots, accompanied by the appearance of a low-dotted class, would occur if the two Ac2's were situated in the same chromosome but not in juxtaposition and were separable by crossing over. The frequency of the low-dotted class would depend on the amount of crossing over. If testcross ratios are the sole criterion, it is not possible to differentiate between two linked Activators with 50% recombination and two which are unlinked. The possibility that the infrequent low-dotted kernels found in the testcross progeny of plants with two linked Ac2's arise not from crossing over between the two Ac2 loci but represent changes in "state" can and will be answered by following the closely linked ms8 gene.
The great majority of the kernels from the reciprocal cross of bz2-m/ bz2-m ac2/ac2 females by bz2-m/bz2-m Ac2/Ac2 males have a stable bronze phenotype since they possess endosperms with one dose of Ac2. However, a low and varying percentage exhibit Bz dots ranging discontinuously in number from one to more than 200 per kernel. Upon testing, most of the kernels with low numbers of dots proved to be heterozygous for two Ac2's while kernels with a high number of dots generally were heterozygous for three, four or even five Ac2 loci. If the different Activators were randomly inherited, ascertainment of genotype was relatively precise and straightforward, but linked Activators complicated the analysis. Despite this difficulty, progress has been made. Plants have been found with three Ac2 loci which assort independently and others where two of the three Ac2's are linked and the third is unlinked.
Exceptional kernels with a very high mutation frequency have given rise to plants heterozygous for three, four and five independent Ac2 genes. A number of testcrossed ears have ratios suggesting heterozygosity for two or three independent Ac2's and two that are linked but an exact genotypic determination calls for additional testcrossing. As might be anticipated from an exponential dosage effect, the number of dots in endosperms with six or more Ac2's is often so great that the pigment from closely spaced dots coalesces to form what appears to the unaided eye as self-colored tissue.
The production of male and female gametes with as many as four or five Ac2's by a plant known to be homozygous for a single Ac2 gene indicates that there may be repeated replications (amplification) of the DNA comprising the Ac2 locus in one mitotic cycle or else there is a stepwise increase in Ac2 number during a series of mitoses. Regardless of the mechanism by which the multiple copies of Ac2 arise, they can be inserted at various positions in the parental chromosome or transposed to heterologous chromosomes. The two alternatives occur in approximately equal frequencies in those instances where the change is from one to two Ac2's (12 cases of two linked Ac2's:13 cases of two independent Ac2's).
M. M. Rhoades and Ellen Dempsey
Return to the MNL 56 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
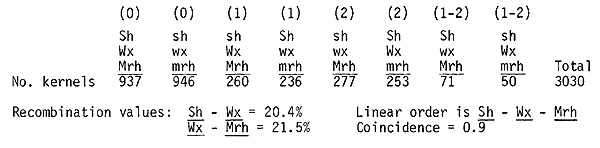{kind=link}
{kind=link}