In our 1981 Maize News Letter report we stated that an abnormal chromosome 10 (K10) terminally deficient for W2 and Sr2 possessed the O7 locus. This gene had been placed in normal chromosome 10 (N10 between the W2 and Sr2 loci and the linear order in the distal end of 10L was believed to be R - W2 - O7 - Sr2. Our data suggest that the linear order in the distal tip of abnormal 10 is R - O7 - W2 - Sr2, i.e., the O7 through W2 segment is inverted in abnormal 10 relative to its orientation in normal 10. However, we questioned if the putative order in N10 is correct, since it was largely based on F2 data from plants with different genetic backgrounds and recombination values are known to be influenced by genetic modifiers. We now present evidence that the tentative linear order in the distal tip of 10L is indeed correct and that the segment including W2, O7, and L13 genes, which lies between R and Sr2 in N10, is inverted in abnormal 10.
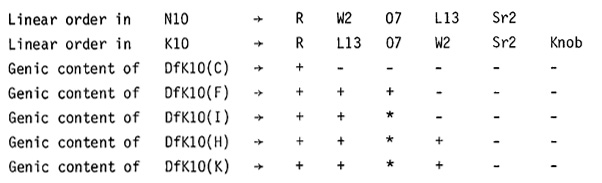
The deficient abnormal chromosome 10 mentioned above was one of 11 deficient abnormal chromosomes 10 that we isolated in our high-loss studies. They all arose following rupture of a dicentric bridge arising at the second microspore mitosis as a consequence of the delayed replication of knob heterochromatin. Delayed (incomplete) knob replication occurs in those microspores with two or more B chromosomes. Five of the deficient abnormal chromosomes 10 had terminal deficiencies for the distal end of the long arm of 10. They were not involved in any structural rearrangement. The remaining six deficient abnormal chromosomes 10 possessed a translocated segment from a heterologous chromosome. Most of our attention has been given to the five simple terminal deficiencies, which were designated as DfK10(C), DfK10(F), DfK10(H), DfK10(I), and DfK10(K). All five were tested for possession of specific loci by ascertaining the phenotype of DfK10/N10 compounds where N10 carried a recessive mutant allele. If the hemizygous individuals expressed the mutant trait, the tested DfK10 chromosome lacked the wild type allele present in the parental K10 chromosome. Our findings are summarized in the following table where the + sign indicates that the tested Df chromosome had the wild type allele, a - sign denotes a lack of it and an * signifies that tests have not been completed. The listing of the tested loci at the top of the table is their linear order in N10.
All five of the DfK10 chromosomes lack the K10 knob and the Sr2 locus, since the loss of the dominant Sr2 allele present in K10 was used in screening for DfK10 chromosomes. The relative lengths at pachynema of N10, K10, and the five K10 chromosomes with simple terminal deficiencies are given below:
In DfK10(C)/N10 heterozygotes the two chromosomes 10 are of the same length at pachynema. The differential segment with its three small knobs allows recognition of the DfK10 chromosome. Despite having a length equal to that of N10, it is deficient for all tested loci distal to R, indicating that these missing genes are situated in the euchromatic region of K10 located between the differential segment and the K10 knob. Cytogenetic studies of all the DfK10 isolates are in agreement with the conclusion that this segment of euchromatin distal to the differential segment is homologous in gene content to the terminal 0.3 of N10 where the linear order is W2 O7 L13 Sr2. However, the linear order of the W2 O7 L13 region is inverted in K10 where the linear sequence is R L13 O7 W2 Sr2 knob (the inverted stretch of chromatin is underlined). This conclusion, which we reported in 1981 based on the uncertain location of O7, has now been confirmed by employing the L13 mutant which was shown by Peter Mascia to lie five crossover units proximal to Sr2.
DfK10(C) and DfK10(F) are the most instructive since their constitution for all extant loci in distal 10L has been determined. DfK10(F), cytologically a shorter deficiency than DfK10(C), is deficient for the K10 knob, the Sr2 and W2 loci, but not for O7 or L13. DfK10(C) lacks all of these marker genes. It should be, and cytologically is, a longer deficiency than DfK10(F). It should be recalled that the DfK10 isolates all arose in high-loss plants in which faulty knob replication at the second microspore mitosis, induced by B chromosomes, results in dicentric bridges at anaphase. They are ruptured. The telophase nuclei become transformed into sperm cells. A sperm with a broken end produced by bridge breakage may unite with the egg pronucleus to form the zygote. It is known from McClintock's earlier work that healing of freshly broken ends occurs in the zygote and they subsequently behave as stable ends. There is no breakage-fusion-bridge cycle, which can lead to structural changes such as duplications. That is, we recover unchanged in the zygote the immediate products of bridge breakage in the ultimate division of the male gametophyte.
We first considered the possibility that even though the portion of K10 carrying the W2 O7 L13 segment was transposed distally relative to its position in N10, the linear order with respect to R and Sr2 remained the same. However, if this were true, all breaks in the dicentric bridge between O7 and W2 would yield DfK10 chromosomes deficient for Sr2, L13, and O7 but not for W2. This was not the case since Df(F) has the L13 and O7 genes but not W2 or Sr2. Such a chromosome could not arise if the order were normal. With an uninverted order, all breaks between R and W2 result in chromosomes lacking W2 O7 and L13, but DfK10(F) is not deficient for O7 or L13. The order of the W2, O7, and L13 genes is clearly inverted in the distal euchromatic segment of K10 which is homologous to the terminal 0.3 of the long arm of N10. The linear order is R L13 O7 W2 Sr2. Only the R locus occupies the same position in the two homologues. An alternative order of R L13 W2 07 Sr2 in K10 can be ruled out since a single break in the dicentric bridge between L13 and W2 would produce a DfK10 chromosome missing the W2, O7 and Sr2 loci. DfK10(F) is deficient for W2 and Sr2 but not for O7. Our studies with the six DfK10 isolates involving translocations are not as extensive as those with the terminal deficiencies but they are in no way in disagreement with the conclusions drawn from the simple deficiencies.
Although DfK10(F) and DfK10(I) are both deficient for W2 and Sr2 but not for L13, they are not identical. They differ in pollen transmission. DfK10(F) is frequently male transmitted, producing in F2 progenies kernels homozygous for Df(F). These kernels have an aleurone layer mosaic for colored-white spots and give rise to albino seedlings. Df(I) is rarely, if ever, transmitted through the pollen. The two deficiencies also differ in the amount of recombination between the R locus and the tip of the deficient chromosome in DfK10(F)/K10 and DfK10(I)/K10 compounds. In testcrosses of DfK10(F) R - / K10 r Sr2 heterozygotes there was 15% recombination between R and the end of the Df(F) chromosome while in DfK10(I) R -/K10 r Sr2 compounds there was less than half as much crossing over.
Of some interest is the fact that the frequency of crossing over between R and the tip of the deficient chromosome in DfK10(C)/K10 heterozygotes is ca. five percent and most of the exchanges take place in the homozygous differential segment with its three small knobs. The extent of this segment in DfK10(C)/K10 meiocytes, in which crossing over distal to R can occur, is physically as long as the R-Sr2 interval in N10 homozygotes where there is 36% recombination. One is forced to conclude that recombination is low per unit length of chromatin in the differential segment. Unfortunately, this chromatin is not populated with mutant loci and its origin is unknown.
DfK10(K) has the shortest deficiency. It lacks only Sr2 and is pollen transmissible but the amount of recombination between R and its tip in DfK10(K)/K10 heterozygotes is only 19 percent. This interval is approximately twice as long at pachynema as the R-Sr2 region in N10/N10 bivalents where the recombination value is 36 percent. Either the large heterozygous K10 knob is depressing crossing over in regions adjacent to it or there is a high exchange frequency in the tip of normal 10L, where none can occur in DfK10(K)/K10 individuals since it is in a hemizygous condition. This question is being resolved.
Apparently some misunderstanding exists regarding the architecture of abnormal 10 found in races of maize. All of the abnormal chromosomes 10 observed by Kato in different races of maize had the same structure in that the segment in 10L with the three small knobs (our differential segment) was not adjacent to the K10 knob but was separated from the knob by a stretch of euchromatin. In K10/N10 pachytene bivalents, the differential segment is paired with the distal 0.3 of the long arm of N10. Kato found that some races of teosinte have an abnormal 10 with the differential segment next to the knob while others possess the maize type. The two kinds of abnormal 10 present in teosinte may differ by a single paracentric inversion but this has not been demonstrated to be so. Since teosinte crosses readily with maize to give fertile hybrids it is not obvious why the second type of abnormal 10 found in certain teosintes has not undergone introgression into sympatric races of maize.
M. M. Rhoades and Ellen Dempsey
Return to the MNL 56 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}