At least 31 independently discovered sources of cytoplasmic male sterility have been classified by one or more means into one of three types: C, S, and T (Beckett, Crop Sci. 11:724, 1971; Gracen and Grogan, Agron. J. 66:654, 1974; Forde et al., Genetics 95:443, 1980; Kemble et al., Genetics 95:451, 1980). None of the 84 sources mentioned by Duvick (Adv. Genet. 13:1, 1965) was known to have arisen as a mutation within a homogeneous population of N (nonsterile) cytoplasm plants. We were interested, therefore, in characterizing two additional sources of cytoplasmic male sterility that had been discovered in China.
One source (designated Chi31cms) was identified in 1974 as one of several male sterile plants in a population designated Luo Synthetic 1 originally derived from 10 USA lines and 2 China lines. From 1974 to 1976 this source was maintained by crosses with fertile plants of Luo Synthetic 1. In 1977 backcrossing with the inbred, Chi31, was begun and our material was analyzed after the third backcross.
The second source (designated Mo17cms) was found in 1976 as a single male sterile plant within a population of the inbred line C103 which, according to records in China, was not uniform. This line was advanced by crosses with Mo17 from 1976 through 1979.
In 1980 these two male-sterile lines and their respective maintainer lines were grown at St. Paul, MN. We inoculated leaves of 5-6 week-old plants with pathotoxin from Helminthosporium maydis race T. Lesions developed on leaves of the Mo17cms line and on Wf9T check plants but not on any other lines. This suggested that Mo17cms belonged to the T cytoplasm group. We also crossed the two sterile lines with Wf9 lines carrying nuclear genes for fertility restoration of T, S, and C cytoplasm as indicated in Table 1.
Fertile progeny from Mo17cms were obtained only in crosses with the T cytoplasm restorer line. The 1 fertile:1 sterile segregation indicated that Mo17cms was homozygous dominant at either the Rf1, or Rf2 locus. Wf9S Rf3/rf3 restored fertility to all the progeny in crosses to Chi31cms as expected if only pollen carrying the Rf3 allele were functional. We have no explanation for the 3 fertile plants in progeny from Wf9T tester crosses. These fertility restoration data indicated that Mo17cms belonged to the T class and Chi31cms belonged to the S class.
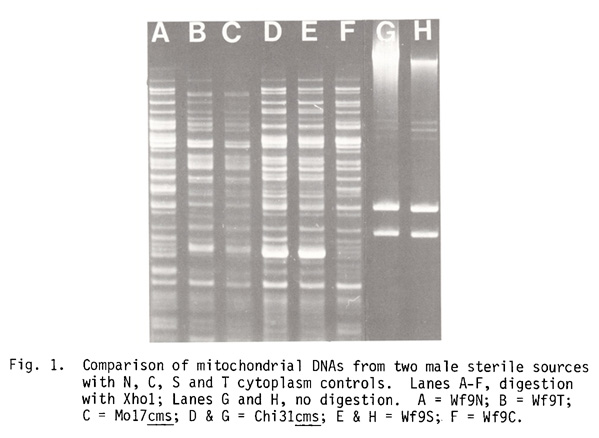
We confirmed this classification by comparing mitochondrial DNA restriction endonuclease digestion patterns of the 2 sources with those of Wf9N, C, S and T cytoplasm controls (Fig. 1). The Xho1 restriction pattern of Mo17cms (lane C) was indistinguishable from that of Wf9T (lane B) and the Chi31cms (lane G) and Wf9S (lane H) patterns both clearly show the S1 and S2 plasmid-like DNAs characteristic of the S cytoplasm group (Pring et al., PNAS 74:2904, 1977).
Records on the identity and sources of the original lines comprising Luo Synthetic 1 are no longer available but it is possible that one line could have been S cytoplasm with restorer genes. Subsequent segregation of rf3 alleles could have uncovered the S cytoplasm male sterility. S cytoplasm has not been used in China for hybrid production. It is also possible that the T cytoplasm source in C103 resulted from a seed mixture some time prior to 1976. Therefore, it is very likely that Chi31cms and Mo17cms trace back to other S and T sources, respectively, and not to new independent events. It remains of interest, however, to determine whether the same cytoplasmic event(s) leading to male sterility can occur repeatedly and independently.
Xie You-ju* and Burle Gengenbach
*Department of Agronomy, Peking Agricultural University, The People's Republic of China
Return to the MNL 56 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}