Two cytosolic superoxide dismutases, designated SOD-2 and SOD-4, have been reported in maize (Baum and Scandalios, Differentiation 13:133, 1979). These two proteins have been purified to homogeneity and partially characterized (Baum and Scandalios, Arch. Biochem. Biophys. 206:249, 1981). SOD-2 and SOD-4 are virtually indistinguishable on the basis of a number of biochemical properties including their sensitivities to various inhibitors such as KCN, NaN3, H2O2 and diethyldithiocarbamate. In addition, the two isozymes are recognized identically by antibodies raised against SOD-4. However, the nuclear gene Sod4 affects SOD-4 expression but has no obvious effect on SOD-2 expression, suggesting that SOD-2 and SOD-4 may be encoded in distinct structural genes (Baum and Scandalios, J. Hered. in press, 1982). This study has been extended to include an amino acid analysis of the two superoxide dismutases.
Duplicate lyophilized samples of SOD-2 and SOD-4 were hydrolyzed in 0.7 ml 6N HCl under nitrogen in 1 ml vials (Pierce Chemical Co.) at 145 C for 2, 4 and 8 hr. The contents were neutralized and dried in a sodium hydroxide vacuum dessicator and analyzed with a Durrum 500 amino acid analyzer. The labile amino acids, threonine and serine, were estimated by extrapolation to zero-time hydrolysis. For the other amino acids, the estimates obtained for the 2, 4 and 8 h hydrolysis periods were averaged together because the recoveries of each amino acid were statistically the same for each sample (± 10%). SDS-polyacrylamide gel electrophoresis was performed on a 13.5% gel according to the method of Laemmli (Laemmli, Nature 222:680, 1970).
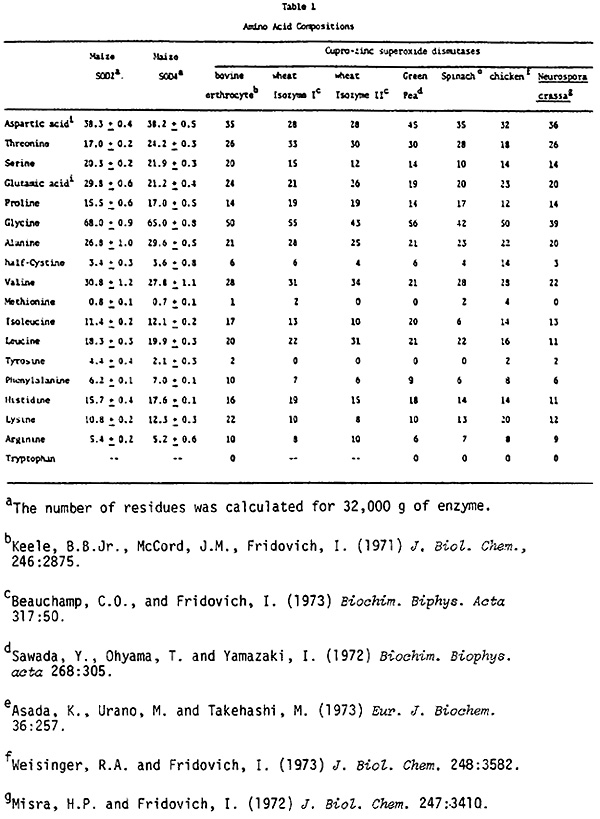
The amino acid compositions of the two proteins are shown in Table 1 along with amino acid composition data for several copper and zinc-containing superoxide dismutases purified from other eukaryotes. SOD-2 and SOD-4 differ considerably in their glutamic acid, threonine, and tyrosine content, indicating that the two isozymes are distinctly different proteins. Similarities among all of the superoxide dismutases in Table 1 can be observed, reinforcing the notion that they may be conserved in their structure. More definitive information on the evolution of these proteins could be obtained through determination of their amino acid sequences.
Although no attempt was made to determine tryptophan, it is clear that copper and zinc-containing superoxide dismutases are generally deficient in this amino acid. The higher content of glutamic acid in SOD-2 could account for the difference in electrophoretic mobility observed between the two proteins on non-denaturing gels as well as for their chromatographic behavior on anion-exchange columns.
J. A. Baum and J. G. Scandalios
Return to the MNL 56 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}