The enzyme UDPglucose:flavonoid 3-0-glucosyl transferase (UFGT) catalyzes the 3-0-glucosylation of flavonols and anthocyanidins (Larson, R. L., Phytochemistry 10:3073, 1971; Larson, R. L. and Lonergan, C. M., Planta 103:361, 1972). Studies on stable and unstable recessive mutants of the bronze locus have provided evidence that UFGT is encoded at this locus (Larson, R. L., and Coe, E. H., Biochem. Genet. 15:153, 1977; Dooner, H. K., and Nelson, O. E., Biochem. Genet. 15:509, 1977, and PNAS 74:5623, 1977). A partial purification scheme for UFGT has been described (Dooner, H. K., and Nelson, O. E., PNAS 74:5623, 1977). We have prepared highly purified UFGT by DEAE-cellulose, CM-cellulose, Affi-Gel Blue and hydroxylapatite chromatography. A simple assay and the purification scheme are briefly described here.
The assay is carried out in a buffer containing 25 mM Tris (pH 8.0), 10 mM CaCl 1mM dithiothreitol (DTT), 2 mM UDPG, 1.25 µCi/ml C14 UDPG (240 mCi/mmole, Amersham): 25 µg/ml human or bovine serum albumin, and 2 mM quercetin (dissolved at 40 mM in EGME). The enzyme is assayed in a volume of 20 µl, adding 1-5 µl of the enzyme to a suitably concentrated reaction mixture. After incubation at 37 C for 30 min, the reaction mixture is extracted with 100 µl of n-amyl alcohol by vortexing; the phases are separated by a brief centrifugation in a Beckman Microfuge and the alcohol phase is removed to a double GF/A glass fiber filter (Whatman). The filter is dried and counted in a liquid scintillation spectrometer. When the labeled products appearing in the alcohol phase are analyzed by paper chromatography, the major labeled component is quercetin 3-0-glucoside; small amounts of labeled material having a different Rf are observed when crude extracts of mature kernels are assayed, but these are no longer detected after chromatography of the enzyme on DEAE-cellulose.
Large-scale preparations are made from mature Bz kernels. Dry kernels are pulverized to a fine powder using a high speed flour mill (Magic Mill II, available from Magic Mill, 235 West 200 South, Salt Lake City, Utah 84101). The powder is extracted in the cold with 3 ml/g powder of 0.1 M NaCl, 0.05 M Tris-HCl (pH 7.5), 1 mM DTT containing 20 mg/ml AG 1-X2 anion exchange resin (Bio-Rad) to adsorb pigments. Extraction is carried out for 1 hr with gentle stirring. The extract is centrifuged at 10,000 rpm for 20 min at 2-4 C in a Sorvall GSA rotor. The supernatant is filtered through siliconized glass wool. The pellet is resuspended in 1/3 of the original volume of buffer by brief homogenization in a Waring blender, respun and the supernatants are combined and dialyzed overnight in the cold against 0.01 M NaCl, 0.01 M Tris HCl (pH 7.5), 1 mM DTT, 25% glycerol, then centrifuged at 10,000 rpm for 20 min at 2-4 C. The supernatant is again filtered through glass wool and loaded on a short, large-diameter DEAE-cellulose column (2 ml bed volume/g starting material), equilibrated with 0.01 M NaCl, 0.01 M Tris-HCl (pH7.5), 1 mM DTT, 10% glycerol. The column is washed with 3 column volumes of 0.1 M NaCl, 0.01 M Tris HCl (pH 7.5), 1 mM DTT, 10% glycerol and then eluted with an 0.1-0.2 M NaCl gradient in the same buffer. The enzyme begins to elute at about 0.15 M NaCl.
Active fractions from the DEAE-cellulose column are pooled and dialyzed overnight against 0.08 M NaCl, 0.02 M MES (pH 5.0), 1 mM DTT, 50% glycerol and applied to a CM-cellulose column equilibrated with the same buffer containing 10% glycerol. The column contains 0.5-1.0 ml packed resin/g starting material. The column is washed with equilibration buffer and the material flowing through the column is assayed for enzymatic activity. More than 90% of the total protein eluting from DEAE-cellulose with UFGT adsorbs to the CM-cellulose column at this pH, but the UFGT enzyme does not. The pooled active fractions are dialyzed against 0.01 M NaCl, 0.01 M Tris-HCl (pH7.5), 1 mM DTT, 50% glycerol and applied to an Affi-Gel Blue (Bio-Rad) column, using 1-2 ml packed bed volume/100 g starting material. The column is equilibrated with the same buffer containing 10% glycerol. After loading, the column is washed with 3 column volumes of 0.1 M NaCl, 0.01 M Tris-HCl (pH 7.5), 1 mM DTT, 10% glycerol and eluted with an 0.1-0.5 NaCl gradient in the same buffer. The pooled active fractions from this column are dialyzed against 4 mM Na2HP04 (pH 7.0), 1 mM DTT, 50% glycerol and applied to a hydroxylapatite column (Bio-Rad HTP) equilibrated with 4 mM Na2HPO4 (pH 7.0), 1 mM DTT, 10% glycerol, using 1 g packed bed volume/200 g starting material. The column is eluted with a 4-10 mM phosphate gradient containing 1 mM DTT and 10% glycerol. The pooled active fractions are dialyzed against 0.01 M Tris HCl (pH 7.5), 0.01 M NaCl, 1 mM DTT, 50% glycerol and stored at -20 C.
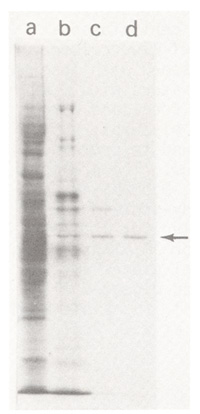
All procedures are carried out in the cold. The order of the CM-cellulose and Affi-Gel Blue columns can be reversed. The amount of protein contained in the active fractions becomes difficult to measure directly by UV absorbance beyond the CM-cellulose chromatography step and active fractions are generally examined by polyacrylamide gel electrophoresis to monitor purity. The proteins co-eluting with the enzymatic activity from DEAE-cellulose, CM-cellulose, Affi-Gel Blue and hydroxylapatite columns are displayed on a Coomassie Blue-stained SDS polyacrylamide gel in Figure 1. The arrow indicates the 50 kD protein identified as UFGT. The enzyme becomes increasingly unstable with purification and the assays should be done in the presence of serum albumin after the DEAE-cellulose step. Enzyme purified through DEAE-cellulose chromatography is stable for many months in 0.01 M Tris HCl (pH 7.5), 0.01 M NaCl, 1 mM DTT, 50% glycerol at -20 C. Highly purified enzyme can be stored frozen in liquid nitrogen in the same buffer. However, the effect of repeated freezing and thawing has not been determined. The purified protein from mature tissue migrates as a closely-spaced doublet of the same apparent molecular weight on 2-dimensional isoelectric focusing and SDS-polyacrylamide gels. Only one of the two spots is present in immature aleurone tissue. We do not know whether the charge heterogeneity is introduced during tissue dehydration and storage or during purification.
N. Fedoroff and J. Mauvais
Return to the MNL 56 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}