The exact nature of the mutator system (Mu) is not known at the present time. Its transmission does not follow traditional Mendelian patterns. When mutator plants are outcrossed generation after generation most of the outcross progeny have mutator ability. If mutator is confined to the chromosomes at one or more defined loci there must be some mechanism for selecting Mu bearing chromosomes for transmission. Evidence bearing on such a system of selective transmission has been negative, to date. In any case, this model would predict that by intercrossing Mu plants, the mutation frequency might be increased by twofold, but an increase above twofold would not be expected. Mutator could be chromosomal and increases in mutation frequencies above twofold could be found if Mu were transposable in nature and thus capable of inserting copies of itself into new chromosomal locations. The normal outcrossing procedure for propagation of Mu would tend to hold the number of transposable elements more or less constant because with each outcross the haploid mutator genome is being combined with a virgin genome into which Mu elements can be inserted. On intercrossing (i.e., inbreeding) Mu lines, however, the number of Mu elements would begin to accumulate. After several generations of inbreeding there could indeed be many Mu elements distributed over the genome, resulting in a mutation frequency considerably higher than twofold.
If Mu is transmitted extrachromosomally (it is transmitted with equal facility through the male and female) it might well build up in numbers upon inbreeding, resulting in mutation frequencies greater than two times those observed in Mu outcrosses.
To determine if inbreeding resulted in a twofold or greater increase in mutation frequency, a series of crosses was made.
The simplest and most efficient method of inbreeding is to self-pollinate for several generations. This is not practical with Mu stocks because progeny of selfs consist of many weak, runty, deformed plants (mutants?). Things do not improve with additional generations of selfing. To avoid these difficulties crosses were made between two different Mu outcrosses. Lines were chosen that produced two ears, one for selfing and one for crossing. The selfs permit the elimination of any plants that carry a mutation induced in one or more of its parental gametes. The protocol followed in this experiment is outlined in Figure 1. In the first generation, four different families of Mu outcrosses were planted and crosses made between individual plants of these four families. The male parent in each cross was self-pollinated, crossed to the female Mu plant and outcrossed to a standard line. The second ear of the female Mu parent was selfed and the plant was also outcrossed to a standard line. The parent plants are termed Mu1 plants and their outcrosses are Mul outcrosses. The F1 of the cross between the two Mu lines was called Mu2 per se. Fifty or more seeds of each outcross were planted, the resulting plants self-pollinated and a sample of seeds from each ear was tested for the presence of seedling mutants. The same procedure was followed for the Mu2 per se cross. The next generation two Mu2 plants were crossed and at the same time outcrosses to standards were made. The product of Mu2 X Mu2 was termed Mu4 per se and the outcrosses Mu2 outcrosses. Next, Mu4 plants were crossed and outcrossed to give Mu8 per se and Mu4 outcrosses, and lastly, Mu8 plants were crossed to give Mu16 per se, and outcrossed to produce Mu8 outcrosses.
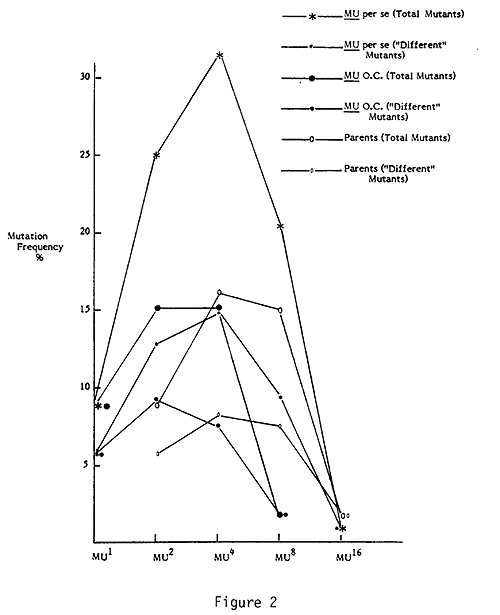
The results are plotted in Figure 2.
When Mu2 (= Mu x Mu) plants are produced, the male Mu parents contribute to the Mu2 plants their mutant bearing pollen grains in a frequency expected of single Mu plants. The female parent also contributes to the Mu2 plants, through the ovules, a frequency of mutants expected of single Mu plants. Thus, the Mu2 per se plants should have twice the frequency of mutation that the Mu1 parents have. The Mu2 mutant frequency, however, is greater than expected. Instead of two times the Mu1 frequency it is 2.26 and 2.84 times. For the latter value, the difference between the calculated two times Mu1 rate and Mu2 is close to being significant at the 5% level (X2 = 3.0936; p 5%, X2 = 3.841). Since both the Mu1 parents were tested as males the greater than expected increase in mutation frequency observed in Mu2 plants would be expected if more mutants were transmitted through the female Mu1 parent. However, previous tests of transmission of mutants in reciprocal crosses of Mu plants revealed significantly fewer mutants transmitted through the female. There are at least three possible explanations for the greater than twofold increase: 1) there is something about Mu sperm nuclei traversing Mu female tissue that results in additional mutants being produced in the sperm; 2) the Mu sperm and Mu egg nuclei in a common embryo sac may result in additional mutants being produced in either or both of them; or 3) in the zygote the presence of two Mu genomes may induce some additional mutations.
When Mu2 plants are used in outcrosses, they do not have two times the mutator activity as might be expected for either of the models dealing with the nature of the mutator system suggested earlier in this report. The mutation frequency of Mu2 outcrosses does not approach that of the Mu2 per se progeny. For the total mutant values the difference is very significant. Thus, combining two Mu's in a single individual produces less of an effect than combining the products of two separate Mu individuals. The effect of two Mu's combined in a single individual is not additive. Each Mu under these conditions seems to be less effective than when present in separate plants. In other words, the two Mu's together are more effective than a single Mu but not twice as effective as a single Mu. Thus it is reasonable to assume that Mu4 per se would show another increase in mutation frequency, and indeed it does.
Mu4 outcrosses on the other hand, show virtually no change in mutation frequency when compared to Mu2 outcrosses. Thus it appears that four doses of Mu is either not adding anything more to the system or the increases in number of Mu's may be resulting in their partial inactivation. Inactivation is suggested because Mu8 per se is observed to have a statistically very significantly lower rate than Mu4 per se. When Mu8 is outcrossed, there is observed very low mutation activity, in spite of the fact that Mu8 per se had a mutation frequency over twice that of Mu1. Eight doses of Mu seems to be too much for the normal functioning of Mu's. Almost all Mu activity seems to be lost. This loss of activity is reflected in Mu16 per se, which has a mutation frequency slightly less than the Mu8 outcrosses.
Inactivation of Mu was suggested above as an explanation for the loss of mutator activity in these plants with putative high doses of Mu. If it is indeed an inactivation phenomenon due, perhaps, to high concentrations of Mu, then outcrossing for several generations of the Mu16 per se plants to non-Mu lines or continued outcrossing of Mu8 outcrosses, again for several generations, may restore high mutability to these lines. Restoration of mutator activity under these conditions would only occur if the Mu inactivation is reversible. The loss of activity may be due to permanent inactivation of Mu or perhaps an actual destruction of Mu. In either case, it would not be possible to restore Mu activity by outcrossing.
There are other possible explanations. Perhaps cells with high doses of Mu can not compete. If there is some mitotic sorting out mechanism for Mu such as occurs for plastids in variegated plants, then the cells with few or no Mu's would out-compete those loaded with Mu and give rise to an essentially non-Mu- plant. Yet it is difficult to envision why such sorting out would take the Mu activity down to such a low level (to about 1%) when Mu plants with 5-9 percent mutation frequency do not seem to have their vigor affected. Such a precipitous drop in only one generation would not seem likely on the basis of this model.
In discussions of these results with Dr. Hugo Dooner, he suggested that sterility may be involved. Higher doses of Mu may be causing abortive pollen and/or ovules, leaving only low dose Mu or zero Mu ovules and/or pollen to function. I did not follow pollen sterility in these experiments. We have had occasion, however, to screen large numbers (several hundred) of Mu1 plants for pollen sterility and a lesser number (about 50) of Mu2 per se plants and in neither instance were many plants with abortive pollen found. An occasional plant with lower sterility (35% or less abortive pollen) did occur and a very rare semi-sterile plant was found.
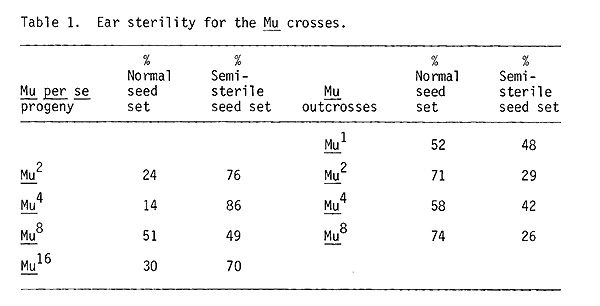
I did classify the ears for semi-sterile seed set throughout this experiment (Table 1). Seed set is not a very good gauge of actual sterility because it is so easily influenced by environmental factors, particularly in more inbred stocks such as the Mu per se lines. These lines did indeed have a high frequency of ears with poor seed set. But note that Mu1 outcross lines also have high incidences of ear sterility although pollen examination has established that Mu1 outcrosses show very little pollen abortion. There does not seem to be any consistent pattern of increasing sterility at higher Mu doses in either the per se crosses or the outcrosses expected if high doses of Mu were inducing sterility.
Donald S. Robertson
Return to the MNL 56 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}