Preliminary evidence has suggested that resistance to the maize disease caused by Cochliobolus carbonum race 3 is characterized by a reduced rate of disease increase and is quantitatively inherited. This study was designed to determine the nature of gene action governing resistance in maize to this disease.
Three sets of diallel crosses were made. In each of the first 2 sets, 7 maize inbred lines with different levels of resistance were crossed in all possible combinations. Three inbreds from each of these crosses were used as parents for the third cross. A total of 134 F1 and selfed progenies were screened for resistance to the pathogen in the growth chamber when seedlings were approximately 3 weeks old. Assessments for resistance were based on disease efficiency (DE), lesion length (LL), and sporulation capacity (SC).
The underlying genetic model in the diallel analysis assumes diploid segregation, no reciprocal differences between the progeny families, homozygous parents, no genotype-environment interactions, 2 alleles per locus, and absence of non-allelic gene interactions as well as linkage. The variance within each array of the diallel table and the parent-offspring covariance were calculated for each of the 3 replications by averaging the values of the F1's including reciprocals. The difference between the parent-offspring covariance and array variance was calculated for each replicate and averaged over the 3 replicates. Scaling tests were carried out to determine if the assumptions made for the diallel analysis were met. Regression of the parent-offspring covariances on array variances were computed to determine an appropriate model for the 3 characters in the study.
Analyses of variance of the diallel tables, similar to that of K. Mather and J. L. Jinks (1971, Biometrical Genetics), were computed to test the significance of the main genetic components, and to detect the presence of reciprocal differences among the progeny families.
The difference in parent-offspring covariance and array variance in crosses involving 2 parents in the one diallel were not homogenous. Therefore, they were removed from the analysis. The results of regression analyses of parent-offspring covariances on array variances indicated that a simple additive-dominance model with additive environmental effects and absence of gene interaction is adequate to describe the data. Only additive effects of genes were of primary importance in the inheritance of DE, LL, and SC. Average heterosis, line heterosis, specific combining ability effects, and maternal effects were occasionally significant, but were always much smaller than additive effect.
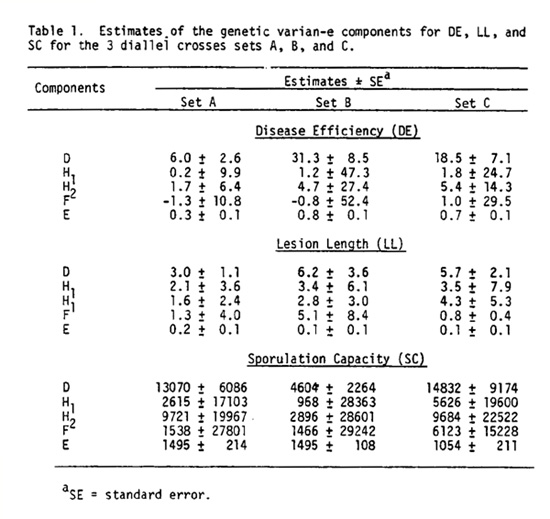
The only genetic variance estimate that exceeded twice its standard error was D, the additive genetic effect (Table 1). Other components H1 (estimates of the dominance effects), H2 (dominance effects measuring asymmetry of positive and negative effects of genes), and F (an indication of the covariance of additive and dominance effects) were smaller than their own standard error estimate and thus assumed not to be important. Environmental component of variation was symbolized by E.
The narrow-sense heritability estimates for DE, LL, and SC were 0.83, 0.60, and 0.37 respectively, indicating that it should be possible to select for resistance with reasonable success.
A. H. Hamid, J. E. Ayers and R. R. Hill, Jr.
Return to the MNL 55 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}