Investigations of the nuclei in the developing endosperm have been initiated. The ultimate goals of these studies are (1) to elucidate the cytological and molecular behavior of this tissue, and (2) to determine the feasibility of using this system for in situ hybridization. Mitotic index calculations, nuclear volume measurements, microspectrophotometer measurements of DNA, and cytological observations were made from A188 endosperms every two days following pollination. In addition, tritiated thymidine, chemical pretreatments, and in situ hybridization studies were performed using several different strains.
Generally, the 3N endosperm nucleus begins dividing within hours after fusion, and continues dividing rapidly for several days. At about the third or fourth day after fertilization, cell wall formation takes place. By the fifth day, the endosperm is entirely cellular. Cell and nuclear divisions eventually cease in the central region of the endosperm, and persist the longest time in the peripheral regions. The nuclei then become very large in the innermost region. Considerable starch formation can be noted by the twelfth post-pollination day. Any nuclear division during these later stages is found only in the outer aleurone region.
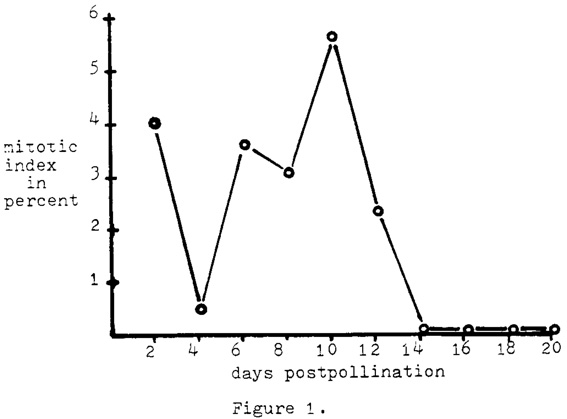
Mitotic index was determined for every two-day period of post-pollination from A188 selfs by making propionic carmine squashes of endosperm tissue fixed in Farmer's solution. The tissue was always taken from the central region of the endosperm. Random transects of the slide preparations were made, and all endosperm nuclei encountered in the microscopic field were tabulated. From 194 to 1150 cells were counted for each two days of post-pollination through day 20 (Fig. 1). Not a single endosperm nucleus was observed in mitosis after post-pollination day 12. It is not known if the data for day 2 and day 4 are biologically significant or simply the results of aberrant samples. The collection of data at these stages will be repeated.
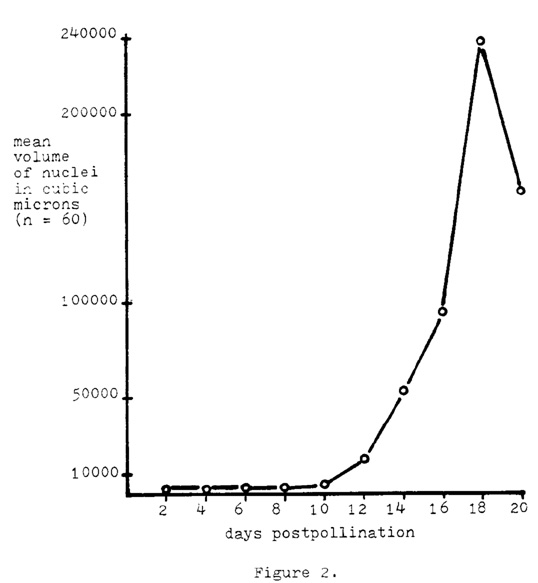
Nuclear volumes in cubic microns were obtained from propionic carmine preparations which were made in the same manner as described above. Care was taken so as to prevent manual pressure to the nuclei, other than the placement of the cover glass. The nuclei in the endosperm tissue from A188 selfs were measured for every 2 days of post-pollination through day 20. Measurements were made with a Zeiss multiple operations processor MOP-3 system, and its units were subsequently converted to micron units. Random transects were again made, and all whole nuclei encountered in the microscopic field were measured until n equalled 60 at each stage. The diameter was determined by measuring the nucleus in both the longest and shortest directions and then averaging the two figures. Volumes were calculated assuming a spherical shape. A large range in nuclear size was noted within the tissue at each of the post-pollinated stages, especially within the older dates. The mean nuclear volume at two post-pollination days was 937.0 cubic microns, and at 18 post-pollination days the mean was 230,195.0 cubic microns (Fig. 2). This constitutes a mean volume increase of 255-fold, with some individual nuclei increasing as much as 975-fold. These increases also constitute a mean growth rate of 10.3 cubic microns per minute during the time period between day 2 and day 18. The decrease in mean volume at day 20 appears to be due to a nuclear breakdown that begins to occur in these older stages, with the larger nuclei being the most vulnerable. Generally, the nuclei after about 12 post-pollination days do not have a surrounding cytoplasm; rather they exist as naked entities. A marked increase in nuclear volume occurs after about 10 days. The mean growth rate from day 10 to day 18 was determined to be 20.3 cubic microns per minute.
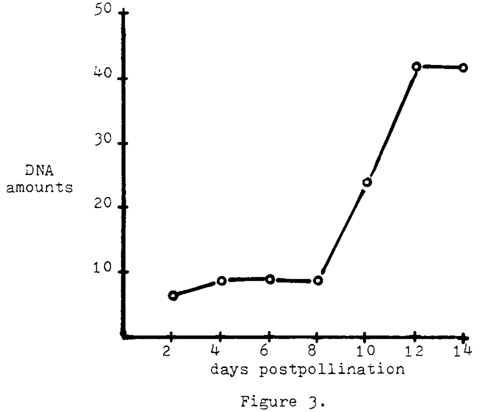
Means for the DNA amounts per nucleus were determined for each two-day post-pollination period from day 2 through day 14. Feulgen technique was applied to the nuclei and the two-wavelength method was used with a Zeiss microspectrophotometer to gain the DNA measurements. A total of 30 nuclei were measured at each post-pollination stage. A mean increase in DNA amounts of 6.8-fold was calculated for the period between day 2 and day 14 (Fig. 3). A very sharp incline was observed after day 8.
The following linear correlation coefficients have been calculated from
these data:
| Nuclear volumes | DNA amounts | |
| Days post-pollination | .829 | .906 |
| Nuclear volumes | -- | .730 |
All of the linear correlation coefficients are highly significant, statistically. In addition, a multiple correlation among all three variables shows a .906 correlation coefficient which is also highly significant. In this calculation, relative DNA amounts served as the dependent variable, and the nuclear volumes and days post-pollination were the independent variables.
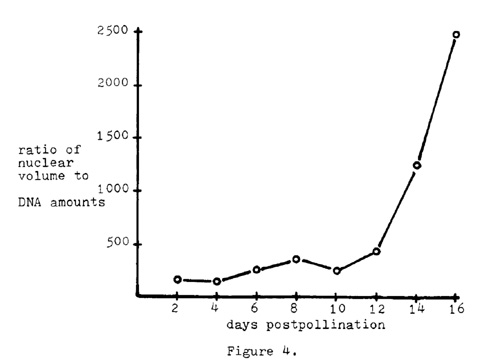
Regardless of these high correlation coefficients, it can be noted that the nuclear volume increases at a much higher rate than does the DNA amount per nucleus. Figure 4 plots the ratio of nuclear volume to DNA amounts against days post-pollination.
These data prompt several conclusions and several more speculations. In the first place, there seems to be little question that the amount of DNA per nucleus increases as the endosperm develops. Feulgen photometry shows this DNA increase occurring somewhat concomitantly with nuclear volume increases. In addition, no mitoses are observed after 12 post-pollination days. In conjunction with this latter observation, tritiated thymidine autoradiographic experiments resulted in silver grain production directly over the chromatin of the nuclei, at stages later than 12 days. It appears that polyteny, endopolyploidy, or some other form of gene amplification is occurring. Since the DNA increase from day 2 to day 14 is approximately only seven-fold, the possibility exists that only certain regions of the chromatin complement are amplified. This amplification of a limited number of areas, however, may have to be immense to reach the seven-fold DNA increase that has been noted. Such conditions, if they do exist, may be ideal for in situ hybridization analyses of genes that are unique, or with few copies, in normal diploid tissue.
In situ hybridization with 125I-labeled rRNA was successfully accomplished with these nuclei. A major drawback is the inability to recognize specific chromosomes because of the very diffuse state of the chromatin. Sixteen different chemical pretreatments at two different temperatures have not alleviated this problem.
Additional experimentation is presently being carried out and planned (1) to further elucidate the genetic system of the endosperm nuclei, and (2) to determine the feasibility of the system for in situ hybridization experiments. In this regard, cytological analyses, additional pretreatments, the use of cytological markers such as B chromosome translocations, and more autoradiographic studies are in progress. (Supported in part by NSF Grant PCM-7912069 A01 and administered through the University of Minnesota.)
Richard V. Kowles, Ronald L. Phillips and A. S. Wang
Return to the MNL 55 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}