In our screening of more than 150 embryos of the inbred normal line of maize BP, it was possible to isolate 2 wholly insensitive to cumulative LT 2.5 mM growth inhibition. The embryos were submitted to two inhibitory culture cycles, 10 days each, and then were transplanted to an aerated sterilized organic culture medium without asepsis. The first inhibitory cycle was carried out in total darkness at 30 C, and the second was carried out with a photoperiod of approximately 12 hours, also at 30 C. One of the isolated embryos (mutant?) died in the second inhibitory culture cycle, and the other died after 10 days of culturing in the organic unaseptic medium. This is another case in which the desirable isolated plant is not capable of completing its whole life cycle. The results obtained by us were less successful than those of Hibberd et al. (Planta, 148:183, 1980), since these authors used the isolated callus technique and then by tissue redifferentiation were able to obtain developing, although infertile, plants.
It is evident that in addition to the selective (selective?) effect the LT method causes several unexpected effects that alter normal plant metabolism. We call the selective effect the primary LT effect, and the metabolic alterations the secondary LT effects.
In our case, the BP line has more than 30 years of inbreeding, is a stable line, and no apparent mutations have been found (Magoja, personal communication). The isolated possible mutants were morphologically different from "normal" counterparts; both possible mutants had root and shoot lengths similar to normal counterparts growing in control mediums (without amino acids); both had long coleoptiles; both were thinner than normal counterparts and less green, and none of them showed the typical swollen roots caused by LT inhibition.
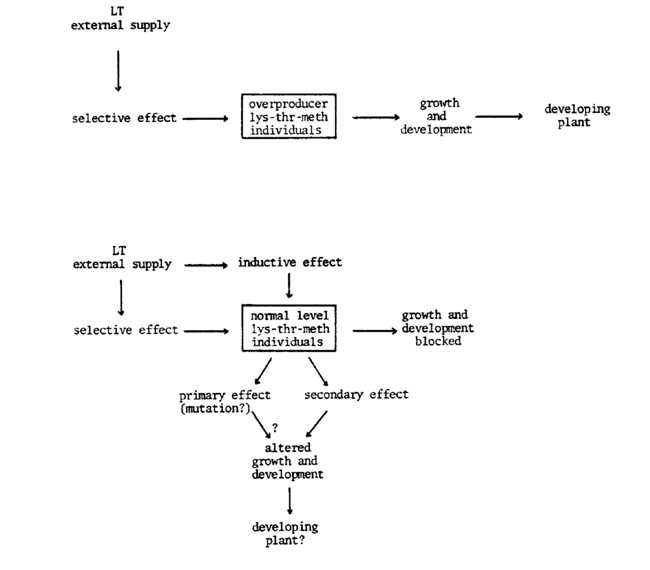
So, it is clear that these isolated seedlings were not BP seedlings with less sensitivity to the LT inhibition and they could have been possible BP mutants. Thus, the striking point to discuss is how these possible mutants appeared provided the mutational stability of the BP line. The only logical answer is to think that an external effect was responsible for the differences and not that the differences (genetic or not) could appear in the presence of the external effect. In other words, we suggest that in certain cases the LT effect is an inductive means of aspartate derived amino acid overproducer individuals rather than a means to select them. The term inductive is not used here as synonymous to mutagenic in the strict sense of the word. The inductive effect (= LT primary growth inhibitory effect) would be the result of an "external pressure" that could affect certain genomal places or produce epigenetic changes.
Obviously, this is not the case for the natural overproducers of essential amino acids, because such individuals do not show inhibition with the LT effect and this method is only selective on them. But the most exciting fact is that these individuals grow normally to physiological maturity (Rapela, unpublished results). A whole view of the problem can be seen as an induction-selection model:
The inductive LT hypothesis does not necessarily involve that the secondary effect always escorts the primary effect. Although up to now there is no evidence of the dissociation between primary and secondary effects, there is sufficient evidence that secondary effects do not act in the same way on all genetic backgrounds (compare Hibberd's results and this). So, the successful possibility would be in the search of a genetic material stable to the secondary LT effect but inducible by the primary LT effect.
Miguel Angel Rapela
Return to the MNL 55 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}