Two-gene control has been suggested previously from analysis of GDH spectra in scutella of maize seedlings (MNL 53:45, 1979). The products of two loci, by free association into hexamers, would produce the seven-band spectrum with binomial staining distribution. However, the latter was found only in developing and mature anthers, while all the other organ-specific GDH patterns deviated widely from the binomial prototype. We therefore suggest that the organ-specific GDH spectra are determined both by differential activity of two genes and by post-translational modification of gene products changing the activities of the associates.
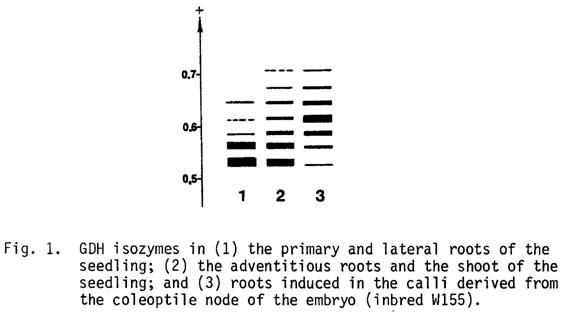
The five-band GDH pattern in the coleorhiza and in the primary and lateral roots of the seedling differ widely from the seven-band pattern in the adventitious roots similar to that of the shoot (Fig. 1, lanes 1 and 2). The adventitious (seminal) roots arise near the top of the scutellar node (Sass, Corn and Corn Improvement, 2nd ed., 1977, p. 97); that is, they are shoot-derivatives. It therefore appears that the GDH epigenotype is determined rather by the origin of the initial cells (stem-cell lineage) than by the further fate of the cells derived from these initials.
Calli derived from the developing and mature maize embryos are regarded as homologues of root and shoot meristems: while M. Freeling et al. (Maydica 21: 97, 1976) interpret these calli to be the apical and lateral root and shoot meristem derivatives, R. L. Mott and W. W. Cure (Physiol. Plant. 42:139, 1978) consider these embryo-derived calli to be abnormally growing adventitious roots. Apparently GDH spectra may help to resolve this discrepancy. Two variants of GDH spectra, that is the scutellum- and the shoot-specific patterns, were found in the calli derived from the coleoptile node of embryos (for details of callus initiation and culture see Khavkin et al., Planta 145:245, 1979). While staining distribution in GDH patterns somewhat varied in the successive subcultures of these calli, yet in none of the cases did we observe the five-band GDH pattern characteristic of the primary or lateral roots. The scutellum-derived calli also produced the seven-band GDH patterns intermediate between the scutellum and the shoot spectra by staining distribution.
By lowering 2,4-D concentration in the agar medium, root formation was induced in the calli derived from the coleoptile node of the embryo. Quite unexpectedly, these roots exhibited the seven-band GDH pattern with the binomial staining distribution (Fig. 1, lane 3), that is, the prototype GDH spectrum free of any subsequent modifications.
The embryo-callus-root transformations may apparently provide a hopeful approach to studies both of the somatic heredity as related to the stem-cell concept and of the modification of maize isozymes in the course of cell and tissue differentiation.
T. B. Sukhorzhevskaia and E. E. Khavkin
Return to the MNL 55 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}