|
Dedicated to the memory of Prof. M. I. Khadzhinov (deceased November 21, 1980) |
Isozymes of 6-phosphogluconate dehydrogenase (6PGD): a presumable mode of control
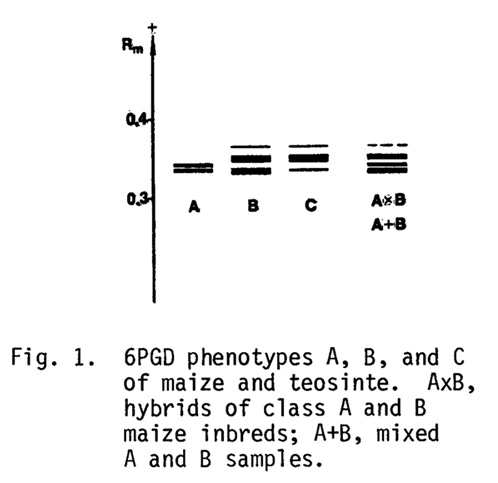
Three phenotypic variants of 6PGD spectra have been previously described in the scutella of 3-day-old maize seedlings (Fig. 1). Most frequently, the pattern consists of two adjacent bands (class A spectrum), while in class B two more distant bands are supplemented with the third weakly staining and more mobile band. Class C spectrum is rather rare: it has been observed in the scutella of Palomero Toluqueño maize as well as in the leaves of all teosinte species, except teosinte Chalco. B and C patterns differ only in their staining distribution (MNL 54:55, 1980).
The probability that the variants of 6PGD patterns are conformers is excluded by the dissociation-association test, by extraction in the presence of increasing 2-mercaptoethanol concentrations, and by separation of isozymes in polyacrylamide gels of different porosity.
To discriminate plastid and cytosol isozymes (allozymes) of 6PGD, we separated a 900 g particulate fraction from the scutella and leaves of seedlings and washed the sediment with hypertonic extraction medium. Twice-washed plastids were found to be microscopically intact and virtually free of alcohol dehydrogenase, which served as a sensitive marker for cytosol impurities. In contrast to the class A pattern, in class B inbreds 6PGD spectra of the cell-free extract (plastids plus cytosol) and the twice-washed particulate fraction (plastids) differed quite distinctly: plastids lacked the most mobile band, while the staining of the second band diminished (Fig. 2). By successive double-dilutions (Klebe, Biochem. Genet. 13:805, 1975) of the particulate fraction extract, the activity of this isozyme was shown to exceed two-fold that of the plastids. We suggest that the class B pattern is formed by such coincidence of two cytosol and two plastid bands that the staining of the more mobile plastid isozyme is intensified by the slower cytosol isozyme. The respective cytosol and plastid bands in the class A pattern apparently do not differ in their mobility (Fig. 2), and therefore two bands are formed in the cell-free extract samples.
Segregation studies (based upon the dimeric structure
of 6PGD, see Gasperi et al., Biochem. Genet. 17:855, 1979) suggest that
classes A and B differ in alleles of a single locus, while the second monomorphic
locus controls the band common both to the A and B patterns:
| Phenotype | |||||
| AA | BB | AB | Chi-square* | P | |
| AAxAA | 10 | - | - | - | |
| BBxBB | - | 10 | - | - | |
| AAxBB | - | - | 25 | - | |
| BBxAA | - | - | 25 | - | |
| ABxAB | 20 | 25 | 40 | 0.877 | 0.50-0.75 |
| BAxBA | 23 | 21 | 46 | 0.133 | 0.90-0.95 |
| ABxAA | 24 | - | 18 | 0.856 | 0.25-0.50 |
| ABxBB | - | 21 | 22 | 0.022 | 0.90 |
| BAxAA | 24 | - | 34 | 1.72 | 0.10-0.25 |
| BAxBB | - | 22 | 22 | - | 0.999 |
*Theoretically expected ratios 1:1:2, 1:1:2, 1:1, 1:1, 1:1, respectively.
Hybrid spectra AxB and BxA are similar to the patterns in A+B mixtures and in AB heterozygotes from the collection of maize races (MNL 54:55, 1980). No additional bands of different mobility due to hybridization of subunits have been observed in F1, F2, and backcrosses among the maize races and teosinte. These data seem to exclude the existence of interallelic heterodimers.
Quite recently, C. W. Stuber and M. M. Goodman (MNL 54:99, 1980) proposed another two-loci model of 6PGD control comprising nine alleles at the Pgd1 locus and three alleles at the Pgd2 locus, and formation of heterodimers between allelic and non-allelic isozymes. To overcome the discrepancies of the two models, we apparently need some additional data that, in particular, would help to interpret the staining distribution in the class B and C patterns.
T. B. Sukhorzhevskaia and E. E. Khavkin
Return to the MNL 55 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}