Through clonal analysis, we have been investigating the destiny of embryonal cells in corn (M. M. Johri and E. H. Coe, MNL 54:80). The basic strategy is to use the anthocyanin markers, eliminate the dominant allele by x-raying and then examine the clones in various parts of the corn plant. Clones originating from the outermost cell layer (L-I) of the embryonic meristem can be easily distinguished. By analyzing the clones, E. H. Coe and M. G. Neuffer (1978, pp. 113ff. in The Clonal Basis of Development, eds. Subtelny and Sussex, Academic Press) have traced the development of nodes 7 through 20, tassel and ear shoot.
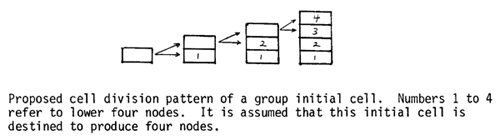
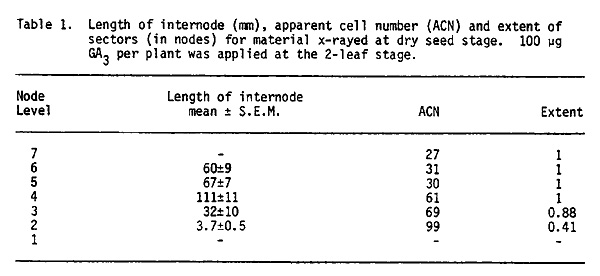
Initials for the first few internodes are present in the dry embryo: In the first six or seven internodes the sectors cannot be distinguished easily because these nodes do not elongate. These nodes were found to elongate upon application of gibberellic acid (100 µg/plant) to the seedling when the second leaf was unfolding. This technique has now enabled us to trace the development of the first few lower internodes also. As shown in Table 1, the sectors occupied only part of an internode for nodes 2 and 3, but a complete internode for nodes 4 through 7.
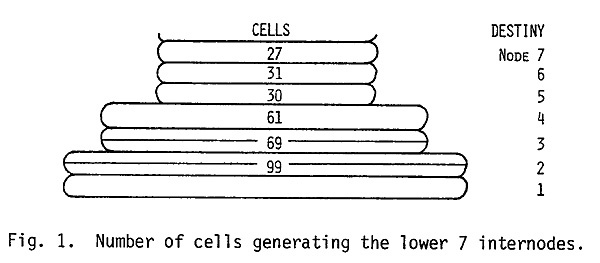
Though no sectors could be observed in the first internode, minute and microscopic sectors were observed in the second internode. These were 4 cells in width and about 16 to 20 cells in length. These data show that, in the meristem of the dry embryo, internodes 2 and 3 are represented by at least 2 layers of cells and internodes 4 through 7 by a single layer of cells (see Fig. 1). These results are also in agreement with the model proposed by Coe and Neuffer (1978). They found that at the dry embryo stage nodes 8 through 14 were represented by three levels of 32 cells each (the 32 cells at a given level developed into 2 or 3 nodes). The data of Table 1 further indicate that determination of individual lower internodes occurs when there are 32 initials in the L-I. These initials divide once generating 64 cells and then the cell divisions leading to the widening and lengthening of the internode axis occur.
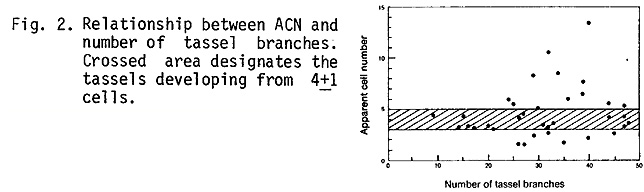
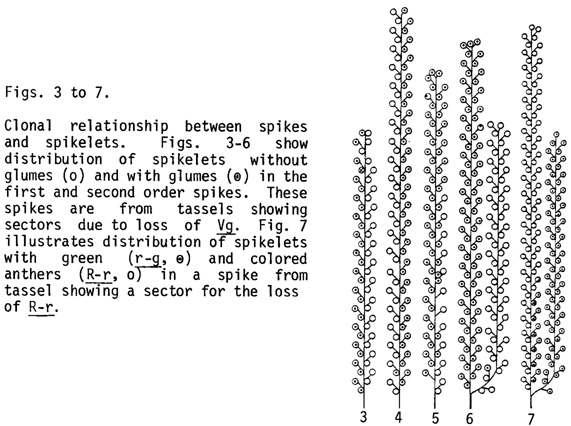
Clonal relationship between spikes and spikelets in the tassel: The tassel sectors described earlier (Johri and Coe, 1980 MNL 54:80) were examined further in order to determine the proliferation pattern of the clones constituting the tassel. Despite considerable variability in results, the size of the tassel seems to be independent of ACN or the number of clones. In the different families investigated about 60% of the tassels developed from 4±1 cells of the dry embryo, and tassels with a small or a large number of branches developed from more or less the same number of embryonic cells (Fig. 2). This observation indicates that in the tassel initials the rate of cell division and the time when spikes get determined are quite different in sparsely branched tassels as compared to highly branched ones. Whether these two processes are coupled is not known at present but these parameters are worthy of consideration in the future if we wish to modify the size of tassel.
The lineages of all four cells are present at the base of tassel, but their relative proliferation is highly variable and finally only two lineages extend into the central spike. The individual first order branches were either completely or partially included in the sector (Figs. 3-5). The latter situation was observed especially in the spikes at the border. The second order spikes were always of one phenotype, completely included in the sector (Figs. 6 and 7). The first order branches can therefore develop from the lineages of two cells but only one of these usually continues into the second order spike. Only rarely one or two spikelets situated at the base of the second order spike were found to have developed from the lineage of a second cell (Fig. 6). The proliferation of a cell lineage in the first order spike is quite ordered and generates a linear file of cells. Often the same lineage developed into several sessile or pedicellate spikelets along one side of a spike. At the border, a single spikelet on a first order branch was occasionally half sectored (Fig. 7), and in such spikelets one floret showed all the three anthers either green (r-g) or colored (R-r); or bronze (bz2) or purple (Pl). A single spikelet can thus develop from two cell lineages but the stamens of a floret develop from a single cell lineage.
Sectoring pattern in ear shoots: The ear sectors observed earlier (Johri and Coe, 1980, MNL 54:80) were studied further and four distinct but related patterns of the proliferation of clones have been found.
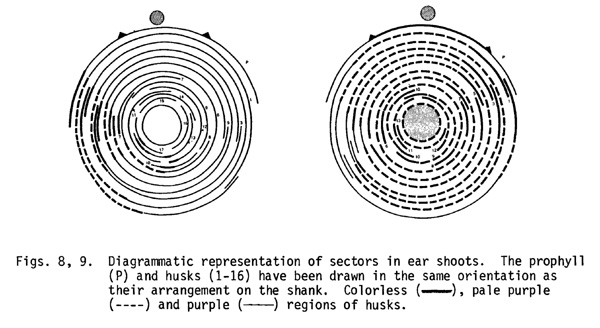
1. In 20% of ears with a sector, the edges of all the husks were observed to be colorless (non-purple). The shank and the husks between the edges were pale purple. The shank lacked any external features to suggest the continuity of colorless edges from one husk to the next. The entire cob was included in the sector. The meristem of these ear shoots can be interpreted as a periclinical chimera in which the L-I layer (or layers) originated from a single L-I cell lacking the dominant allele (B, Pl or C2). The L-II layer in these chimeric meristems was genetically purple. Since we are seeing the underlying purple L-II layers through the colorless L-I cell layers, the shank appears pale purple.
2. In 35% of ears, the sectored region represented between 20 and 90% of an ear shoot. In the ear shown in Fig. 8, the sector consisted of about 25% of the width of the first two husks, then gradually diminished and terminated in the last husk. The sectors in other ears occupied as much as 90% of the ear shoot (Fig. 9) and the progression of the sector from one husk to the next could be easily followed externally on the shank. The L-I in the ear shoots of this class seems to originate from two L-I cells and the extent of proliferation of these cell lineages is highly variable.
3. The third category of sectors was similar to the second one, but showed colorless stripes in the pale purple regions of husks and shank. Some 25% of ears with a sector showed this pattern. These stripes did not follow a clonal relationship from one husk to the next and seemed to represent a displacement of genetically purple L-II by colorless L-I in local areas. Replacement of the genetically colorless L-I cell layers (except the epidermis) by purple L-II cells was also observed and in such cases, purple stripes occurred against a background of pale purple or colorless. The proliferation of two cell layers is thus quite variable and due to periclinal divisions one cell layer can displace the other in local areas.
4. In the remaining 20% of ears, the sector lacked purple color through the depth of the husk. There was no indication that husks in such ears developed from cells derived from both L-I and L-II of the embryonic meristem. Very early during the formation of the ear shoot meristem, one of the initial L-I cells may have divided periclinally so that the L-II was displaced by a L-I daughter cell in the meristem. Alternatively only L-I cells may have formed the bud meristem.
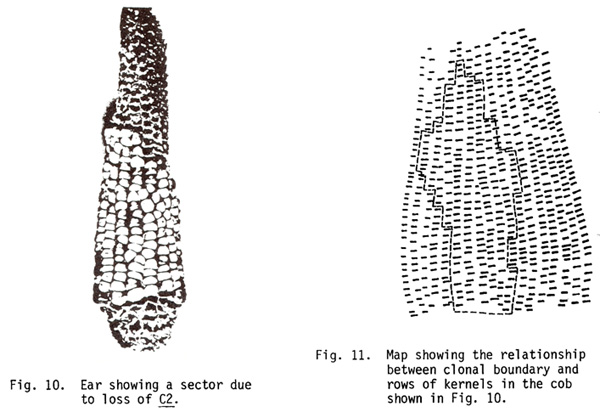
Proliferation of cell lineages in the cob: The clonal boundaries showed no relationship to rows even though a pair of rows develops from a single branch primordium. The clonal tissue switched a complete row or often only half a row laterally as determined on the basis of glume color (Figs. 10, 11). A row of branch primordia does not originate from the lineage of the same cell through the entire length of the cob. Each row can develop from cells derived from two different but adjacent cell lineages. In many instances the florets at the border showed half-sectored glumes.
These results show that the number of times a cell divides is not predetermined. A cell lineage can contribute anywhere between 5 and 90% of husks and/or cob and less frequently only a single cell lineage will extend into a cob. As with the four cell lineages present at the base of the tassel, the lineages of two cells entering the ear are present at the base of the ear. As opposed to the tassel, however, the cob was not divisible into two lateral halves based on clonal restriction. The clonal boundary that runs along the midrib of leaves and through the central spike in the tassel, dividing the corn plant into front and back halves, was not distinguishable in the ear shoot. This seems to be due to a variable proliferation of cell lineages, in which the initials of an ear shoot may be occasionally derived either from the front half or the back half.
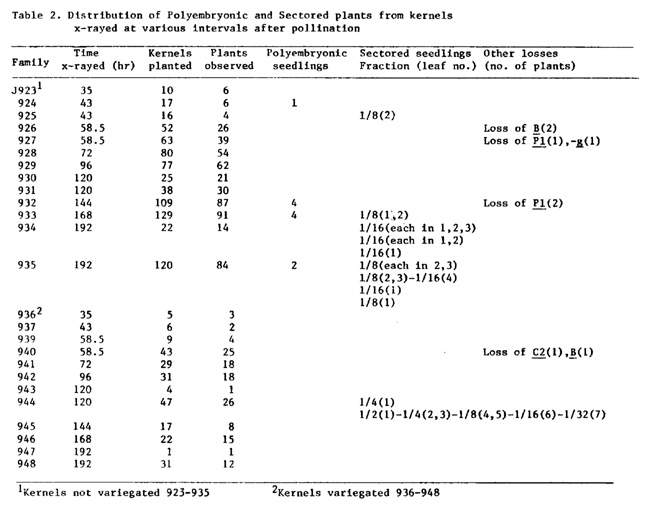
Polyembryonic and sectored plants from embryos x-rayed during early developmental stages: In the first part, we described the results which showed that the initials of the first few internodes were already present in the dry embryos. To determine if these individual nodes were also initially produced as a group, the developing kernels were x-rayed at different stages of maturation. Plants homozygous for anthocyanin and chlorophyll markers (B Pl Wd) were pollinated with wd pollen (male plants were b pl wd with ring-9S carrying C-I Wd). At various intervals (see Table 2) the ear shoots were x-rayed (1000 rad/ear shoot), the plants were returned to the greenhouse and the kernels were allowed to mature. In each family, variegated and unvariegated kernels were sown separately and sectors were scored in the seedlings and mature plants.
Among the different F1 families, about 20 to 60% of the kernels developed into viable seedlings. In the population examined, a few polyembryonic seedlings were also observed. Nine such seedlings arose from kernels x-rayed 43, 144 and 168 hr after pollination. Each of these seedlings exhibited a single coleoptile but two separate and complete axes beginning with the first or second leaf, each having independent sets of leaves and a shoot apex. The two seedlings arising from kernels x-rayed at 192 hr showed a single coleoptile, a single first leaf and a single second leaf. The third leaves were free in one seedling but partially fused in the other.
Polyembryonic seedlings in x-rayed material have been described earlier (D. T. Morgan, 1956, MNL 30:83). Corn embryos show enormous plasticity during development and even the use of x-rayed pollen increases the frequency of polyembryonic seedlings (D. T. Morgan and R. D. Rappleye, J. Hered. 42:90, 1951). The pattern of plasticity suggests to us that possibly the determination of the first and second leaves has already occurred by 192 hr. This can explain the type of polyembryonic seedlings observed in family J935.
The distribution of sectors in leaves strongly supports this interpretation. Most of the seedlings with sectors were present in families J933-935 and J944. White deficiency sectors were present in the first two to seven leaves, suggesting that during development a group of nodes (two to seven) is first produced. In the seedlings of family J935, the sectors were present either only in the first leaf or in leaves two to four. Individual nodes are thus generated starting from the first one (the first node is the lowermost one). Though the data are fragmentary, the most plausible trend indicates that during embryogeny 8 or 16 initials destined to produce a group of nodes are first produced. A single initial cell of this group divides in such a way that it generates a specific nodal initial and an undifferentiated cell. The latter in turn follows the same pattern of cell division and finally all the nodes are formed as shown below:
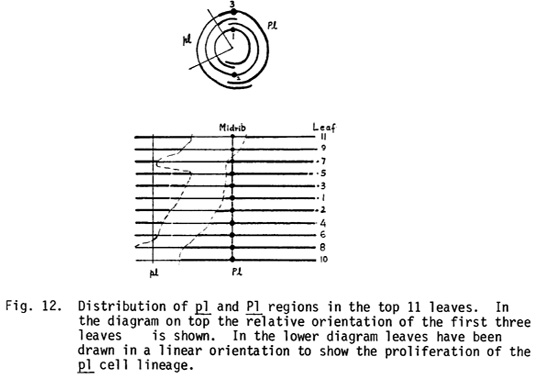
In addition to these plants, sectors representing losses of B, Pl or C2 were also observed. The sectoring pattern of J932-81 will be described. This plant showed a pl sector beginning in the culm below the tassel and continuing up to the 17th leaf from top. It occupied about 25 to 30% of the culm perimeter. The relative distribution of Pl and pl parts as determined on the basis of the sheaths of the top 11 leaves is shown in Fig. 12. At the time of x-raying (144 hr after pollination), there could be three or four cells in the transverse plane in the apical dome of the embryo and one of these presumably lost the Pl marker. The lineage of the pl cell seems to have proliferated in a somewhat random manner. This lineage started left of the midrib in leaves 1 to 7, but was found to be present on the other side of the midrib in leaves 9 and 11.
The first vertical division in the terminal cell of the 2- or 3-celled proembryo has been reported to separate the front and back halves of a corn plant in such a way that the axis of bilateral symmetry is defined by the position of midrib in the leaves (Steffensen, 1968, American J. Bot. 55:354). Our observations suggest that this determination is not absolute and, though the two cells separated by this vertical division usually generate the front and back halves, there is nothing which prevents one cell from contributing lineage to the other side. In the plant under discussion, the lineages of PI cells on the right may have proliferated much less and consequently the lineage of the pl cell from the left of the midrib contributed to the tissues on the right. These observations clearly demonstrate that the proliferation of individual cell lineages is highly variable and spatial location rather than lineal descent is the deciding factor in determining the final fate of a cell.
M. M. Johri* and E. H. Coe
*Permanent address, Molecular Biology Unit, Tata Institute of Research, Bombay, India; current address, Dept. of Biology, Washington Univ., St. Louis, MO 63130
Return to the MNL 55 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}