This project has been described recently (MNL 54:24-26). Briefly, three synthetic populations (A, B, and C) are maintained by random crossing within themselves and have been subjected to x-irradiation (10,000 r to the kernels) for six generations. Synthetic B is the population that will eventually contribute the restructured genome for the allotetraploid and Synthetic D will contribute the normal genome. Synthetic D is composed of 4n material that has never been irradiated. It is derived from 20 maize inbred lines with a large admixture of Alexander's Syn. B. Synthetic A and C are homozygous for a different group of 5 recessive markers: bz2 lg y v16 wx and a su pr gl g. These have been crossed with plants from the B synthetic. Progress toward allotetraploidization will be determined by changes in genetic ratios. This will be reported on next year.
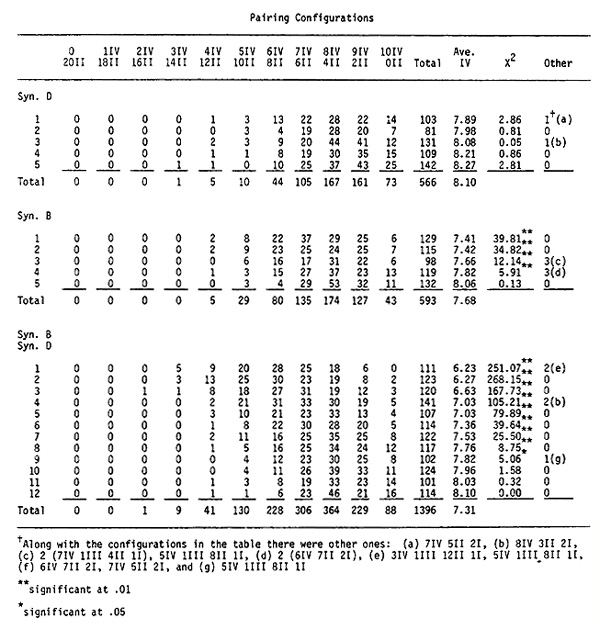
Cytological evidence for allotetraploidization has been found by examining the quadrivalent frequencies of Syn. D, Syn. B, and Syn. BxD plants. The data are given in the accompanying table. The Syn. B plants were derived from the 4th cycle of recurrent radiation. They were self-fertilized once and then crossed with Syn. D plants which were self-fertilized once. The Syn. B plants for the cytological examinations were from the S2 generation.
The pairing configurations were determined at diakinesis. Most configurations were combinations of quadrivalents and bivalents. However, there were a few trivalents and univalents found. These cases were not included in the totals.
The five Syn. D plants had frequencies ranging from 7.89 to 8.27 quadrivalents per cell. There is some variability but it is not significant from the average value of 8.10.
Four out of five Syn. B plants had statistically significant reductions in quadrivalent frequencies as did nine out of twelve Syn. B x Syn. D hybrids. While the declines in quadrivalent frequencies are not great (1.87 quadrivalents at most), the data indicate that allotetraploidization is taking place.
Because the Syn. B plants used were not homozygous the full expression of preferential pairing is not possible. If we symbolize the unaltered chromosomes of Syn. D as S (for standard) and the altered chromosomes of Syn. B as R (for restructured), there are many types of restructuring. Thus there are R1, R2, Rn chromosomes. Many of the chromosomes in Syn. B may not have been altered and thus Syn. B is a mixture of S and R chromosomes. The full expression of preferential pairing is found in balanced ditypes (SSR1R1, SSR2R2 etc.). These would be expected to be rare in the Syn. B or Syn. B x Syn. D hybrids. Most of these plants would be unbalanced ditypes (SSSR1) in which preferential pairing is not possible. Preferential pairing would be possible in tritypes (SSR1R2) but it would be reduced from that found in SSR1R1 or SSR2R2 types. The affinity of R1 for R2 chromosomes may be the same as that of R1 for S or R2 for S. This has been discussed in TAG 54:161-168.
G. G. Doyle
Return to the MNL 55 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}