Polypeptides from primary roots, shoots, pollen and leaf tissue of several maize cultivars were extracted and electrophoretically separated using a modified protocol (see MNL 54, 1980). Primary root and shoot polypeptide extracts were obtained by germinating seeds on moistened filter papers in the dark at 27-28 C for approximately 96 hours, excising 0.5-2.0 g of the terminal 0.5-1.0 cm tips of each tissue into porcelain mortars and homogenizing in two volumes of extraction buffer (200 mM Tris-HCl, pH 7.5, 5% SDS, 7.5% 2-mercaptoethanol and 10 mM phenylmethylsulfonylfluoride). All manipulations were carried out at 0-5 C. The homogenates were transferred to 10 ml glass tissue grinders, further extracted using a motorized homogenizer with a teflon pestle and placed on ice for 5-10 minutes. This last step was repeated and the homogenates were centrifuged in 15 ml corex tubes at 7,700xg for 20 minutes. The supernatants containing the polypeptide extracts were transferred to 5 ml glass tubes, boiled for three minutes and stored frozen at -20 C until required.
For preparation of leaf polypeptide extracts, plants were grown in flats under our standard greenhouse conditions for 21-28 days. After the seedlings were excised at soil level and sliced longitudinally, the young inner leaves were removed and cut up into small pieces prior to homogenization as described for roots and shoots.
For electrophoretic separation of polypeptides 8.3-15% concave-exponential gradient polyacrylamide gels were prepared as described by Laemmli (1970) and the Bio-Rad Gradient Former Instruction Manual (1978). The use of gradient gels resulted in better resolution of a larger number of polypeptides (especially in the 20-100 kilodalton range) than the previously used 8.3% acrylamide, uniform concentration gels.
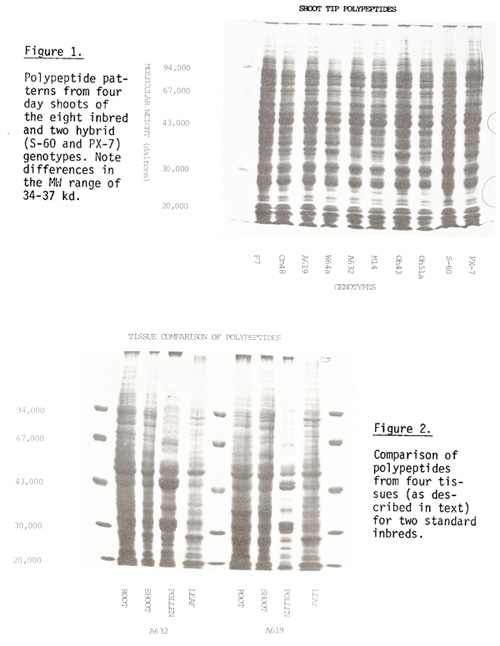
Eight inbreds and two single-cross hybrids were used for electrophoretic comparison of polypeptides among cultivars and between different tissue sources of the same cultivar. In all cases, the overall pattern was similar for any tissue type. Differences in band position and/or intensity were noted primarily in two regions (34 to 37 kd and 80 to 90 kd) for both root and shoot (Fig. 1); prominent bands corresponding approximately to 27, 31, 47 and 51 kd common to all genotypes were present in both tissues. Pollen exhibited differences between cultivars in the bands at 80 to 90 kd (Hughes and Walden, this News Letter).
The polypeptide patterns of the four tissue types were compared for two inbreds (Fig. 2). Root and shoot patterns were similar with at least 85% of the bands apparently being common to both; leaf had 60-70% and pollen had less than 30% of bands in common with those of root or shoot. Bands with molecular weights of 44, 48 and 54 kd were noted in all four tissue types. Although these appear to have the same MW they may represent unique polypeptides in the different tissues. Two-dimensional electrophoretic analysis (currently in progress) should permit this discrimination.
The results indicate that differences may be detected in the electrophoretic phenotypes among cultivars or in various tissues (differing in ploidy level or developmental sequence) within a cultivar. Since culture conditions for any tissue were controlled, the observed differences between cultivars appear to be genotypically determined.
C. L. Baszczynski and W. G. Hughes
Return to the MNL 55 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}