Several laboratories are currently investigating the processing (i.e. posttranslational modification) of chloroplast membrane proteins. One such protein which undergoes processing in maize plastids is the 32,000 dalton lamellar polypeptide. Extensive work in the laboratory of Dr. Lawrence Bogorad suggests the following with respect to the 32,000 dalton lamellar polypeptide in maize: 1) the structural gene for this polypeptide resides on chloroplast DNA; 2) the polypeptide is first synthesized, both in vivo and in vitro, as a precursor with an apparent molecular weight of 34,500 daltons; 3) translation of the precursor takes place on 70S ribosomes; 4) processing of the precursor polypeptide involves a reduction in apparent molecular weight from 34,500 daltons to 32,000 daltons; 5) nuclear information may be necessary for the processing step to occur; and 6) the production of the 34,500 dalton precursor polypeptide is primarily under transcriptional control during photoregulated development (i.e. greening) (for further details see Bedbrook et al. PNAS 75:3060; Grebanier et al. JCB 78:734). Thylakoids purified from the photosynthetic mutant hcf*-3 are missing the 32,000 dalton polypeptide (MGNL 53:38-40); in accordance with (5) above, we expected that this lesion might affect the processing of the precursor polypeptide, possibly resulting in the accumulation of the 34,500 dalton polypeptide and an accompanying loss of the 32,000 dalton species.
In the accompanying figure we present autoradiograms of thylakoid membrane proteins which were labeled with 35S-methionine. Coomassie stained polypeptide profiles are not presented for two reasons: 1) the homology between Coomassie stained bands and radiolabeled polypeptides comigrating with stained bands in the 32,000 to 35,000 dalton region of the gel is uncertain, and 2) the product-precursor relationship described above, as well as the physical mapping data locating the structural gene to chloroplast DNA, have both been elucidated using radiolabeled, rather than stained, polypeptides. For simplicity, the complex pattern of radiolabeled thylakoid membrane proteins obtained upon in vivo or in vitro labeling is not shown in order to focus attention on changes in the 32,000 to 35,000 dalton region of the gel. In control experiments chloroplasts from both hcf*-3 and sib controls were able to incorporate radiolabeled methionine into thylakoid proteins either in vivo or in vitro; the similarity of the patterns obtained from hcf*-3 and sib controls in regions other than the 32,000 to 35,000 dalton region suggests that photosynthetically inactive chloroplasts from hcf*-3 are capable of protein synthesis.
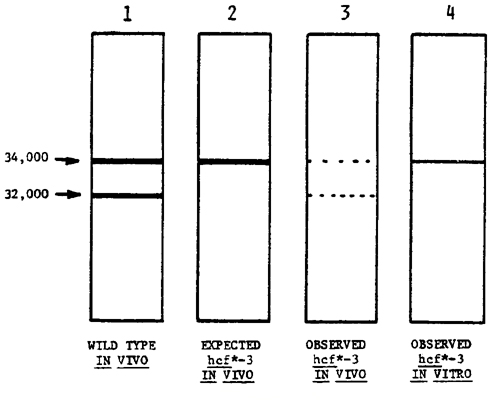
The labeling patterns reported here are in general agreement with the patterns obtained by Grebanier et al.; as shown in lane 1, lamellar polypeptides with apparent molecular weights of 34,000 and 32,000 daltons become heavily labeled when whole leaves are fed 35S-methionine. The small discrepancy between the molecular weight assignments for the larger polypeptide reported by Grebanier (34,500 daltons) and that reported here (34,000 daltons) is probably due to differences in electrophoretic technique and/or gel calibration. According to the data discussed in (5) above, nuclear mutation at the hcf*-3 locus might be expected to block the processing of the radiolabeled 34,000 dalton polypeptide; the anticipated pattern would be as diagrammed in lane 2. In contrast, when whole leaves from hcf*-3 were fed 35S-methionine, very little label was seen at either 34,000 or 32,000 daltons (lane 3), suggesting that the synthesis of the precursor is largely blocked. The small amount of the 34,000 dalton polypeptide labeled in the mutant was accompanied by a similarly small amount of the "processed" 32,000 dalton species.
A further surprising result was found upon in vitro labeling of chloroplasts isolated from hcf*-3; as shown in lane 4, there are indications that these chloroplasts are able to synthesize the 34,000 dalton polypeptide! This would be consistent with the production of a diffusible inhibitor as a consequence of the hcf*-3 mutation; removal of this inhibitor during chloroplast isolation would permit the synthesis of the 34,000 dalton species. Grebanier has reported that isolated plastids are able to synthesize, but not to process, the 34,000 dalton polypeptide; our autoradiograms show little, if any, processing.
These data suggest that the nuclear lesion hcf*-3 may affect transcriptional or translational events in the chloroplast which normally lead to the synthesis of the 34,000 dalton polypeptide; the small amount of this polypeptide present in the mutant is apparently processed normally. The fact that the radiolabeled 32,000 and 34,000 dalton polypeptides are the only polypeptides affected by the mutation suggests that such nuclear control over chloroplast transcription/translation is specific. We cannot as yet rule out (by transmission genetics) the possibility that the chloroplast DNA is specifically altered in plants homozygous recessive for hcf*-3; the absolute lack of photosytem II activity leads to seedling lethality. The in vitro labeling study, however, suggests that the chloroplast DNA is unaltered; production of a labeled 34,000 dalton polypeptide apparently occurs under these conditions. Further experimentation is planned to firmly establish the structural identity of the 32,000 dalton polypeptide produced in vivo with the 34,000 dalton polypeptide produced both in vivo and in vitro; collaborative experiments are planned to distinguish between possibilities of transcriptional vs. translational control.
Kenneth Leto and Charles Arntzen
Return to the MNL 54 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}