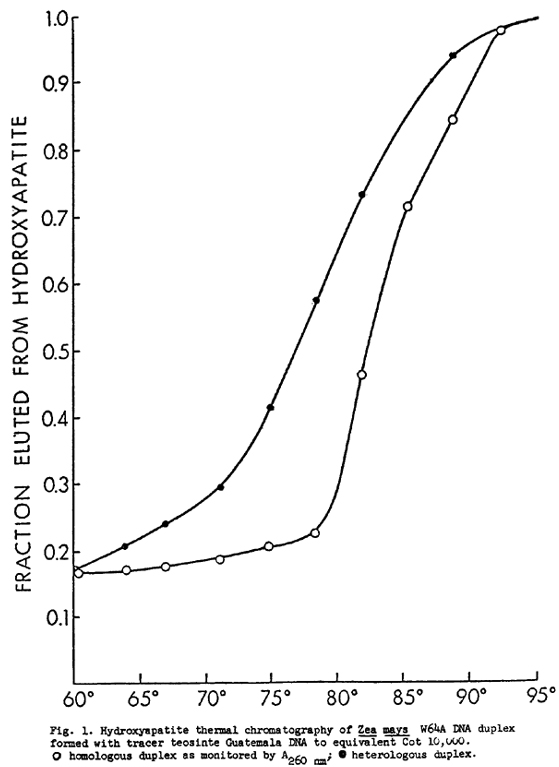
Maize DNA, W64A, was hybridized to DNA of Tripsacum laxus; teosinte, race Chalco; teosinte, race Guatemala; and popcorn, to assess the extent of sequence homology. The divergence between homologous sequences is determined by the thermal stability of hybrid DNA molecules: the difference in Tm, DTm, is approximately equal to the percent mismatch. Maize DNA was reassociated to DNA, 3H-labeled by nick-translation, at a ratio greater than 2000:1; thus, labeled DNA only reassociates with unlabeled driver. The DNA samples were allowed to reassociate to Cot 100 or Cot 10,000, loaded onto hydroxyapatite and thermally eluted in 4-degree increments. The Tm of the homologous duplex, maize-maize, was determined by its A260nm. The Tm of the heterologous duplex was determined by TCA precipitation of chromatographed DNA onto glass fiber filters and counting in scintillation fluid. The data are presented in Table 1. Fig. 1 is a graph of the thermal stability of a Cot 10,000 Guatemala teosinte-maize duplex.
At Cot 100 the reassociated duplexes include most of the repetitive sequences but very few of the unique sequences. The DTms for popcorn, teosintes Chalco and Guatemala are, respectively, 1.7, 2.1, and 1.9 degrees. Maize DNA was also reassociated with labeled self DNA; the DTm was 0.4 degrees. Given the standard deviations, it is difficult to say if one sample is significantly more divergent than another. The extent of reassociation was approximately the same as the homologous duplex in each case. The DTm for Tripsacum is significantly larger, 7.0 degrees, and the extent of reassociation is 56% of the maize-maize duplex.
At Cot 10,000 all repetitive and most unique sequences have reassociated (the Cot-1/2 pure for unique sequences is 2100). The difference in DTm from the Cot 100 results can be attributed to the reassociation of unique sequences. The DTms for popcorn, teosintes Chalco and Guatemala, and Tripsacum are respectively, 2.3, 4.2 3.2 and 8.2 degrees. The percent of shared sequences is approximately equal to the homoduplex in all cases except for Tripsacum which hybridizes to 51% of the extent of the homoduplex. It appears that the unique sequences are all more divergent than the repetitive sequences except for Chalco teosinte. Guatemala teosinte unique sequences appear to be more divergent than Chalco. This difference at the DNA level parallels their morphological characteristics and organellar DNA organization (D. H. Timothy et al., P.N.A.S. 76:4220-4224, 1979). Tripsacum is twice as divergent as teosinte or popcorn from dent corn, but not much more divergent in unique than repetitive sequences.
Table 1. DNA/DNA hybridization of Zea mays to related
grasses
| Cot | Tracera | DTm0Cb | Percent Reassociation of tracer to driver | Number of hybridizations |
| 100 | Zea mays W64A | 0.4 | 100 | 1 |
| 100 | Zea mays Mo17 | 0.95 ± 1.06 | 100 | 2 |
| 100 | teosinte Chalco | 2.1 ± 0.53 | 100 | 2 |
| 100 | teosinte Guatemala | 1.9 ± 1.70 | 100 | 2 |
| 100 | Ladyfinger Pop | 1.7 ± 0.6 | 100 | 2 |
| 100 | Tripsacum | 7.0 | 56 | 1 |
| 100 | wheat | >20 | 10 | 1 |
| 104 | Zea mays Mo17 | 0.2 | 100 | 2 |
| 104 | teosinte Chalco | 2.3 ± 1.0 | 100 | 3 |
| 104 | teosinte Guatemala | 4.2 | 100 | 2 |
| 104 | Ladyfinger Pop | 3.2 ± 0.45 | 100 | 3 |
| 104 | Tripsacum | 8.2 ± 0.64 | 51 | 2 |
| 104 | wheat | > 20 | 10 | 1 |
aDriver in all cases is W64A maize DNA. Teosinte (Z. mexicana) races are identified by region of origin.
bThe DTm is the difference in Tm between driver, monitored by A260 nm, and tracer, monitored by cpm.
Clearly, popcorn and both Guatemala and Chalco teosintes have contributed to the present day maize genome, either ancestrally or due to introgression. Tripsacum is less related in both sequence homology and extent of shared sequences, though there are definite sequences in common. If the changes in the genome occur purely randomly at a clock-like rate of 1% mismatch per 5 million years, we calculate that maize diverged from teosinte 10-20 million years ago and from Tripsacum 40 million years ago. These calculated divergences are unlikely in view of the fact that the grasses are only 70 million years old and individual genera 20-40 million years old. Presumably the intense impact of human selection has contributed to the apparent distance of maize from close relatives.
Sarah Hake and Virginia Walbot
Return to the MNL 54 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}