Structural comparisons of K10 with the B chromosome suggest a direct origin of the B from a complete K10. I have traced Rhoades' photograph of a K10 bivalent from "Mutants of Maize," and darkened the more heavily stained regions of both long and short arms. Beside this tracing is my drawing of a B chromosome (Chromosoma 43:177, 1973). Regions of both the B and K10 have been numbered to facilitate discussion. These chromosomes are remarkably similar from regions 3 to 7, and other differences are minor. Length and chromomere number distinguish region 2. The shorter length of this region in the B chromosome would follow from some condensation as this segment generally appears more heavily stained than euchromatin of the A set. The six chromomeres most usually seen in the B could have arisen by splitting of the three in K10. Perhaps the greatest difference between the drawings is in region 1. However, even here there is little difficulty. The distal euchromatic portion of K10 could heterochromatinize to become the minute short arm of the B. Degeneration of the original centromere and neocentromere formation in the knob would give rise to the new B chromosome centromere. An alternative for the progenitor of the B is KT10 as described by Kikudome (Science 134:1006, 1961). Since no genes other than those dealing with B chromosome function have been found on this chromosome, loss of function of the k10 homologous region would have occurred by heterochromatinization. Because our attention has been focused on the large knob in the long arm of K10, we have been blinded to structural similarity of the B to K10.
Attempts to isolate B chromosome DNA have failed. Studies of DNA extracted from plants of an inbred line with and without B chromosomes could not distinguish B chromatin from the rest of the genome. Analyses of buoyant density and renaturation kinetics, as well as competition studies, led Chilton and McCarthy to conclude that B chromosome DNA was very similar to the remaining genome. These results are precisely those expected if the B arose from an intact K10.
Preliminary observations on C-banded chromosomes add credence to the possibility that B centric heterochromatin is similar to knob chromatin. Although a stock with K10 has not yet been analyzed, other knobs are stained very clearly, either in prometaphase or condensed metaphase root tip chromosomes. Only centric heterochromatin of the B stains heavily in this procedure. No other centric heterochromatin is similarly stained.
The greatest difference between activities of K10 and the B can be found in apparently very different accumulation mechanisms. If the B did arise from K10 we might expect a link between these apparently different systems. I suggest that neocentromere formation may be that link.
B chromosome nondisjunction is seen to be a consequence of delayed replication of the centric heterochromatin and neocentromere formation. Division of the centromere immediately adjacent to the neocentromere could begin pulling chromatids apart only after centric heterochromatin replication, by which time progress to one pole would be too well advanced to prevent nondisjunction.
Also important to consider is why the B chromosome does not undergo preferential segregation during meiosis. Miles has reported K10 derivatives with decreasing knob size that showed progressive reduction in ability to induce preferential segregation. Loss of a sufficient amount of chromatin in the now centric heterochromatin of the B, could therefore render this chromosome incapable of inducing preferential segregation. It is also interesting that KT10 has a reduced knob and lacks preferential segregation capacity.
Proposed mechanisms for B and K10 segregations are consistent with the interpretation of results obtained by Rhoades and Dempsey in their high loss line. In this line, knobbed segments of A chromosomes were eliminated at the second microspore division when two or more B chromosomes were present in the cell. These observations were interpreted as being a consequence of such late replication of knobs that bridges formed at anaphase. Deficient chromosomes and knobbed acentrics resulted from breakage of these bridges. Arguments presented above lead to a slight modification of the hypothesis advanced by Rhoades and Dempsey. Neocentromere formation in the unreplicated knob region would produce a morphologically different bridge; but completion of replication following a break would give the results reported.
A major implication of the suggested origin of B's is homology between these chromosomes and abnormal chromosome 10. Snope could find no supporting evidence in haploids and Ting's report of synaptic homology has been questioned by Rhoades and Dempsey. Loss of synaptic homology could have been an advantage if the B originated in a population segregating the progenitor K10 as a trisomic. Failure to pair with normal chromosome 10 would increase frequency of proper segregation of k1O chromosomes.
If I have stimulated anyone to think of B's and K10 in a different way, this letter will have been worth the effort.
E. J. Ward
(regular address University of Winnipeg, Manitoba, Canada)
Return to the MNL 53 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
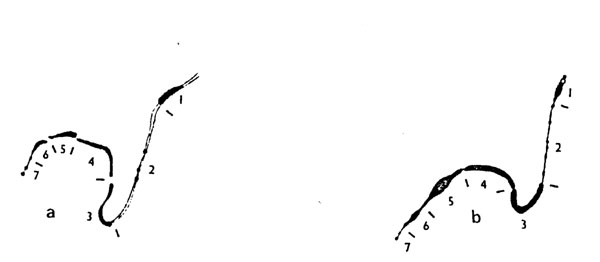{kind=link}