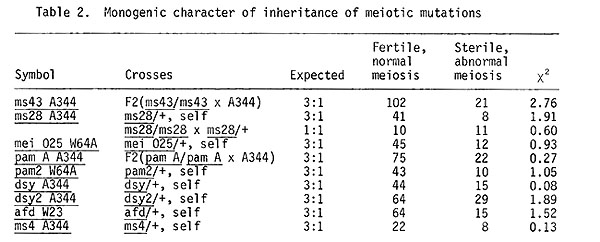
Studies with the identification and analysis of meiotic mutations in maize were started in 1973 as a joint work with Academician M.I. Khadzhinov and his collaborator A. S. Mashnenkov. Initial material was mutations of nuclear male sterility, induced by chemical mutagens in different maize lines (A344, W23, W64A). In all, 52 lines with nuclear male sterility (induced with N-nitroso-N-methyl urea by A. S. Mashnenkov, of P. P. Luk'yanenko Research Institute of Agriculture, Krasnodar) were investigated cytologically by me; in nine of these, meiotic mutations produced sterility. The collected mutations are listed in Table 1 and all mutations are inherited as monogenic recessive mutations (Table 2).
A description of cytological effects of these meiotic mutations on meiosis is given briefly in this communication.
pam A A344, pam2 A344. The pam type of meiotic mutations affected both premeiotic mitoses and meioses. 25% of microsporocytes enter into meiosis as cenocytes. From prophase I, meiosis is desynchronized, so that all meiotic stages from prophase I to tetrads are observed in the same anther. Pycnosis and lysis of chromatin are observed in 18% of the microsporocytes. Various types of meiotic irregularities occur in the other sporocytes (desynapsis, impairment of the spindle apparatus, omission of cytokinesis). Mutant plants show complete male and incomplete female sterility. The occurrence of the pam type mei-mutations indicates that cells are being prepared for meiosis during the last 2-3 premeiotic mitoses (Golubovskaya and Mashnenkov, 1977, Genetika 13:1910-1921).
afd W23. In this mutant the typical steps of the first prophase of meiosis are omitted. At diakinesis, chromosome univalents are randomly scattered in the cells; they are morphologically distinct. At MI, univalents are orderly aligned forming a metaphase plate; at AI chromatids separate to each pole, i.e., centromeres separate during the first division in this mutant; as a result, the second division is much impaired. Gene afd substitutes the first division of meiosis by mitosis.
Studies of such types of meiotic mutations as am and afd open a new approach to the study of genetic mechanisms triggering all the programme from mitosis to meiosis. The existence of an ameiotic mutation blocking meiosis and mutation of afd type indicates that the underlying genetic mechanisms may be quite simple. Such highly specialized cells as meiocytes which have entered meiosis may, as a result of one or two mutation events, return to mitosis.
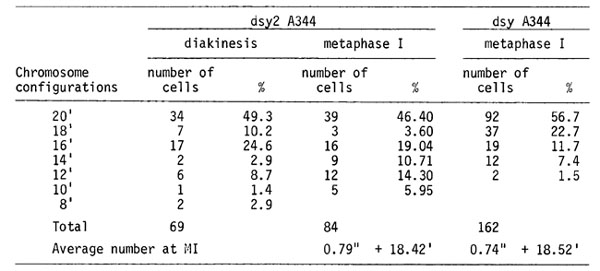
dsy A344, dsy2 A344. Desynaptic mutations are more frequently observed among meiotic mutations. The comparison of chromosome pairing at MI for two of these desynaptic mutations is given in Table 3. At pachytene, regular pairing is disturbed in some chromosome regions. At diakinesis, most chromosomes become univalents. There are, on the average, about 0.8 bivalents per MI cell. The other chromosomes do not pair. The second meiotic division proceeds synchronously in all the cells. The centromeres divide normally at AII. Polyads and tetrads with micronuclei are seen, instead of normal tetrads.
A preliminary electron microscopic study on the desynaptic mutant plants (dsy A344 dsy A344) was performed in the Laboratory of Ultrastructure of our Institute by Christoljubova. It was found that the s.c. in this mutant is abnormal.
To exclude an influence of inbreeding on the structure of the s.c., now we will make a comparison of s.c. in desynaptic plants, segregating from the poor line A344, with ones segregating in F2 from the cross +/dsy A344 (line A344) x +/+ (line W23).
ms43 A344, ms28 A344, mei 025 W64A. These are three meiotic mutations impaired in normal chromosome segregation at the first division of meiosis. The pairing pattern of mei mutant plants homozygous for ms43 is normal. Meiotic irregularities appear only at the first anaphase. The chromosomes are either randomly scattered in the cell (26%), or all the chromosomes remain in its center (33%). The chromosomes of 22% of cells move in groups of ten towards each pole. However, no condensed nuclei arise in this mutant at TI. This very irregular meiosis gives rise to polynuclear pollen grains. Only male sterility is characteristic of these mutant plants.
Meiotic behavior in the mutant ms43 was compared with that of the earlier described maize mutant dv (Clark, Amer. J. Bot. 27:547, 1940). These mutants were found to be similar in meiotic behavior at MI and AI. Their allelic relationships are now under study.
Mutation ms28 A344 impairs the chromosome segregation and moreover blocks cytokinesis partially or fully.
In meiotic mutants mei 025 W64A meiosis proceeds normally until MI. The chromosomes become sticky and cluster at MI and are immobile at AI. Typical TI figures are rarely seen. The chromosomes remain in pycnotic state from MI to interkinesis. Chromatin uncoils at interkinesis, but at MII the chromosomes become sticky again; at AII they are abnormally separated. Cytokinesis proceeds also after the first and second divisions.
Three mutations impairing the different functions of chromosome segregation at the first division of meiosis (ms43 A344, ms28 A344, mei 025 W64A offer an approach to studies of the structure and function of the spindle in the meiotic cells.
ms4 W64A. Meiosis is regular until the tetrad stage in this mutation. The chromosomes of microsporads are subjected to precocious postmeiotic mitosis, chromosomes enter mitosis without replicating and this causes abnormal mitosis. Pollen grains have no pores in mutant plants. It would be needed to compare mei 025 with the mutation po studied by Beadle (Cytologia 5:118, 1933).
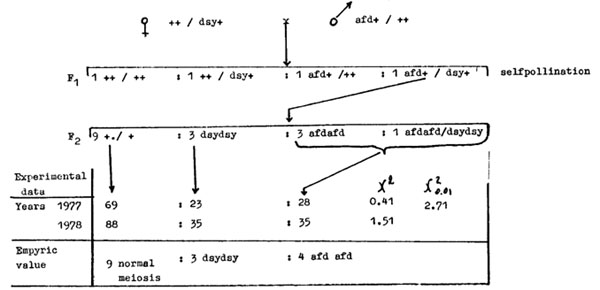
The set of meiotic mutations affecting or blocking different meiotic steps might help to clarify the genetic control of this complex process. The question was to determine whether loci affect the same or different pathways of meiosis. Double meiotic mutants may provide an answer to this question. Two monogenic recessive mutations were chosen as a model; desynaptic mutation dsy A344 and afd W23, substituting the first division meiosis with mitosis. It was possible to obtain a double homozygote for these recessive mutations, because they are completely sterile. A scheme experiment is as follows:
A double heterozygote dsy +/afd + was obtained. The segregation of meiotic pattern was analyzed in offspring obtained by self pollinating this heterozygote. The expected segregation was 9/16 plants with normal meiosis:3/16 plants with dsy type of meiosis:3/16 plants with afd type of meiosis:1/16 plants of genotype dsy dsy afd afd. What will be the meiotic pattern in the double homozygote? If afd and dsy genes control the same sequence of events in meiosis then meiosis in double homozygotes should be either as in the afd type (if afd is earlier involved in meiosis) or of the dsy type (when the desynaptic gene is involved earlier than gene afd). If genes afd and dsy control different alternative pathways of meiosis, then meiosis in a double homozygote should be different from those two mutations.
Analysis of the meiotic pattern in offspring of a double heterozygote demonstrated segregation of 9/16 plants with normal meiosis:3/16 dsy plants:4/16 afd plants (see the scheme above). This means that the genes afd and dsy control the same sequence of meiotic events and that for the accomplishment of events controlled by the gene dsy+, events controlled by the gene afd+ have to take place first.
Concluding I should say that a collection of meiotic mutants can be used:
1. To gain insight into meiosis and its triggering genetic mechanisms; meiosis may be considered as sequential events in a cell and these events are controlled by corresponding genes.
2. For detailed studies on meiotic mutants, which make it possible to establish relationships between cytologically visible structures (the s.c. and chiasmata) and cytogenetic processes (chromosome pairing, crossing over).
3. To establish similarities and differences in processes of meiotic recombination, DNA repair and mutability in eukaryotes.
4. To build models of meiosis occurring in nature. These models help to understand mechanisms blocking single steps of meiosis in apomictic plants and parthenogenetic animals.
I. N. Golubovskaya
Return to the MNL 53 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}