In searching for an explanation of the great morphological and ecological variability of maize, a wide range of hypothetical models have been propounded. Some propose that massive mutation was the major source of new traits, others suggest intergeneric hybridization, while other suggested evolutionary factors are multiple domestications of Zea populations in many places, and long, very effective, divergent breeding by Amerinds. In a new model of maize evolution, facets of each of these play parts, but none is wholly accepted.
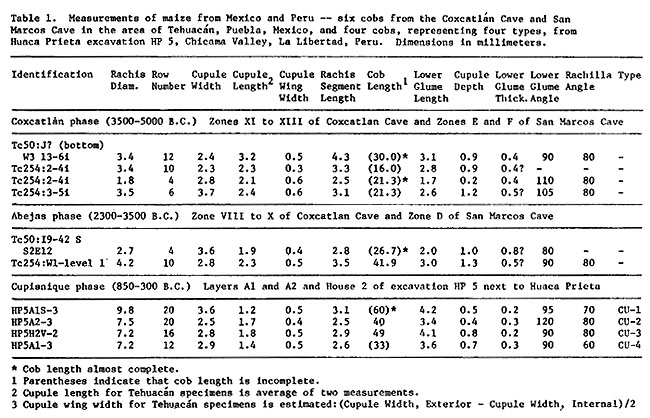
It is proposed that the two principal events in maize evolution are its initial domestication and its hybridization with Zea luxurians (Durieu) Bird, the teosinte of Guatemala and Honduras (R. McK. Bird, 1978. Taxon 27:361-363). Perhaps in central, highland Mexico, a small maize-like plant was domesticated before 5000 B.C., principally by selection for firmer rachillae. Its form could have been much like that of the cultivar found at the lowest levels of Coxcatlan Cave and San Marcos Cave near Tehuacan, Mexico, with short glumes and a narrow, solid rachis (Table 1). Its ca. 64 kernels per ear approximated the number of seed on the fruiting branch of Z. luxurians. (One count of a branch at node 14 below the main tassel gives 65 seed--three to six seed per each of ca. 14 spikes, in cv. Florida grown in Columbia, Missouri by Dr. Jack B. Beckett.)
The cultivar was spread to Guatemala where it eventually hybridized with Z. luxurians. After considerable backcrossing with maize, a subset of the resulting hybrid swarm may have been taken to South America, where because of important new genes or alleles introgressed from teosinte and a new cultural environment, it evolved into a wide range of new races, especially those in the Central Andean Rotund Flour, Altiplano Small Flint and Amazonian Interlocked Flour complexes (R. McK. Bird and M. M. Goodman, 1978, Econ. Bot. 31:471-481). The earliest maize with good provenience on the coast of Peru--from many sites dating 850-300 B.C.--is very different from any Mexican maize yet described (Table 1). Meanwhile, the hybrid swarm in Guatemala, interacting with the original maize and teosinte, produced yet more types of maize through selection in the various cultural environments of Mesoamerica.
As Longley, Kempton and Popenoe suggested in 1937, annual teosinte of Mexico may be another derivative of this hybridization--the great cytogenetic similarity of Zea mexicana and of Mesoamerican-Caribbean Zea mays could be due to their sharing an ancestral stock. Maize containing a relatively high percentage of teosinte alleles could have been moved to Mexico because it had larger ears, kernels with new colors or some other feature. Because two very different species had contributed to this stock, it may have been difficult to stabilize the new maize races. Odd recombinant progeny could have been produced which contained the alleles necessary to the teosinte form, possibly resulting in weedy populations in old corn fields. These may have evolved to the present Zea mexicana, continually affected by introgression from maize (H. G. Wilkes, 1967, Teosinte: the Closest Relative of Maize).
This model does not account for Zea perennis of west Mexico (R. Guzman M., 1978, Phytologia 38:177). Perhaps this also is a product of the hybridization and later movement described above, a segregate which became tetraploid, emphasizing the near-perennial character of Z. luxurians.
Archaeologists may provide evidence critical to testing the model by determining the forms and distributions of pre-5000 B.C. maize and teosinte, the time of first Z. mays-Z. luxurians hybridization, etc. Perhaps they will provide further evidence that people spread teosinte, as seems to be the case with teosinte found in the Tamaulipas caves (P. C. Mangelsdorf, R. S. MacNeish and W. C. Galinat, 1967, Bot. Mus. Leafl., Harvard Univ. 22(2):33-63).
Further morphological and chemical systematic studies are needed, while genetic and cytological studies should provide critical information, especially if they are structured to test the several evolutionary models.
Robert McK. Bird
Return to the MNL 53 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}