In working with selections from the variety Cudu we observed that some selections produced different patterns of pigment formation. Some selections developed only small colored areas at the tip of the kernel while others produced pigment covering the entire crown and perhaps 50% of the surface area of the kernel.
A number of S2 plants selected from Cudu for homozygosity for R-nj and the other required color genes were seeded at staggered dates. A diallel among five early maturing inbreds also was seeded at three different dates in a replicated trial. Several ear shoots near silking were bagged in each row of the diallel set at each planting date. On a given date an ear from each plot was pollinated using pollen from a single R-nj/R-nj plant. Several ears were pollinated from different S2 plants within a 3 day period for each plot. A number of ears were obtained for each row at each planting date which exhibited the navajo color pattern. Colored kernels on each ear were visually rated for proportion of the kernel with anthocyanin pigmentation. Each ear was given a numerical rating. The rating scale ranged from 0 for no color to 9 for completely colored.
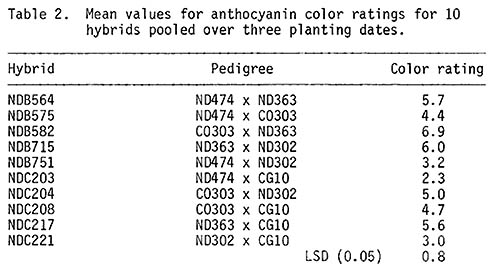
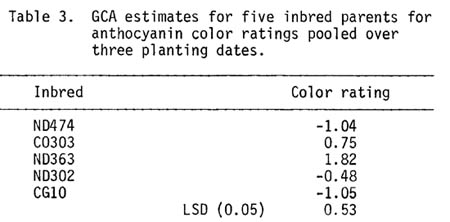
No apparent differences among pollen parents or pollination dates within plots were observed, so all observations within a plot were pooled before computing an analysis of variance (Table 1). The results indicate that differences in color expression among hybrids were due to GCA effects. There were no detectable planting date effects or interactions among planting dates. Color expression ranged from 2.3 for ND474 x CG10 to 6.9 for CO303 x ND363 (Table 2). ND363 had the largest positive GCA effects for color expression while ND474 and CG10 had negative GCA effects (Table 3).
These results seem to contradict those of Kumar and Sarkar (MGCNL 52:119, 1978), who concluded that the variations in the R-nj expression are not heritable and are due to developmental conditions. In this study the differences measured were among maternal parents. If the R-nj anthocyanins are synthesized by plastids as Trautman (MGCNL 49:32, 1975) has proposed for other flavonoids then perhaps interactions among nuclear genes and cytoplasmic factors might account for the differences between these results and those of Kumar and Sarkar. Also the genetic background of the R-nj source was different.
Harold Cross
Return to the MNL 53 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}