As reported previously (MGCNL 51:55-59) screening for families segregating high fluorescent seedlings led to the identification of mutants blocked specifically in photosystem II. Whole leaves and chloroplasts isolated from these photosystem II mutants have been extensively analyzed and the results are tabulated here:
Seedlings were grown in locally constructed growth chambers to the two or early three leaf stage and mutants identified visually on the basis of high levels of chlorophyll fluorescence. Families segregating seedlings originally designated hcf*-19 were found to segregate two types of mutants. One of these mutants was fully blocked in photosystem II activity and emerged as a green seedling (hcf*-19G) while the second type was partially blocked in photosynthesis and emerged as a yellow green seedling (hcf-19YG). Both mutants were typically yellow green at the three leaf stage. The photosystem II mutants therefore fell into two classes, those fully blocked in photosystem II activity (hcf*-3, hcf*-19G) and one mutant which was partially blocked in photosystem II (hcf*-19YG). Chloroplasts isolated from the fully blocked mutants were unable to carry out photosystem II dependent electron transport while the partially blocked mutant hcf*-19YG retained a limited capacity for photosystem II dependent electron transport.
Photosystem II can be roughly divided into two "sides," an electron acceptor side (with unknown component Q as primary acceptor) and a donor side which terminates with a photoactive pigment, P680. In order to determine which side of photosystem II was affected by the mutational blocks signals from the primary acceptor (C-550 and variable fluorescence) and from the photoactive pigment (F695) were monitored. As shown above the acceptor signals were absent (or reduced) in these mutants while the signal from the photoactive reaction center pigments (F695) was present. This suggests that the lesions cause the loss (or inactivation) of the primary acceptor of photosystem II. This loss is only partial in the case of the "leaky" mutant hcf*-19YG.
In algae the mutational loss of photosystem II activity has been correlated with the loss of cytochrome b-559, a component which may play a role in the transfer of electrons between photosystem II and photosystem I. In isolated chloroplasts the cytochrome exists in at least two forms, a high potential form (b-559HP) and a low potential form (b-559LP). Both forms of the cytochrome are greatly reduced or absent in the fully blocked mutants hcf*-3 and hcf*-19G. In a preliminary report (MGCNL 51:57-59) hcf*-3 was described as having lost only the high potential form of the cytochrome; further tests revealed the loss of both forms. Chloroplasts isolated from the partially blocked mutant hcf*-19YG showed a partial loss of the high potential form of the cytochrome and a nearly normal level of the low potential form. These findings are consistent with the observations made earlier in algal photosystem II mutants where the loss of cytochrome b-559 is considered to be a secondary effect of the loss of photosystem II integrity.
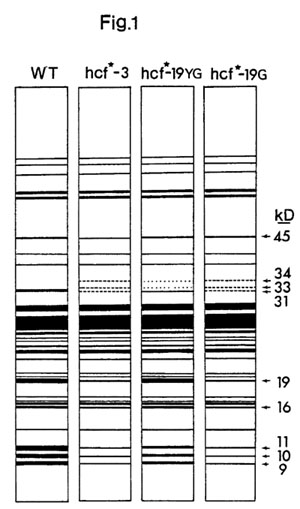
Gradient slab polyacrylamide gel electrophoresis of SDS solubilized lamellar polypeptides suggests that the loss of photosystem II activity is correlated with the loss of a major lamellar polypeptide with an apparent molecular weight of 31kD (kilodaltons; Figure 1). This loss is nearly complete in the case of the fully blocked mutants and is intermediate in extent in the partially blocked mutant hcf*-19YG. This polypeptide is probably not cytochrome b-559 itself, since the molecular weight of the protein moiety of the cytochrome on SDS gels has been reported to be about 5600 daltons. The loss of the 31kD polypeptide is accompanied by the appearance of two minor polypeptides with apparent molecular weights of 33kD and 34kD. Additionally the staining intensity of polypeptides with apparent molecular weights of 45kD, 19kD, 16kD, 11kD, 10kD and 9kD is slightly reduced relative to wild type in the fully blocked mutants.
The major change however is the nearly complete loss of the 31kD polypeptide in the fully blocked mutants. The fact that the amount of this polypeptide present in lamellae correlates well with the extent of photosystem II competence suggests that this polypeptide is in some way necessary for the proper function of photosystem II.
Recently Bedbrook et al. (PNAS 75:3060-3064) suggested that the prominent 32kD polypeptide (most probably our 31kD polypeptide) is synthesized on chloroplast DNA Bam fragment 8 as a 34.5kD precursor which is subsequently cleaved by cytoplasmic products to yield the membrane bound 32kD polypeptide. Possibly then the nuclear mutants reported here are involved in regulating the post transcriptional modification of this protein. The observation that the loss of the 31kD polypeptide is accompanied by the appearance of minor species with slightly higher molecular weights is at least suggestive of a product-precursor relationship (this work was supported by NSF grant PCM 76-08831 to DM).
Kenneth Leto and Donald Miles
Return to the MNL 53 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}