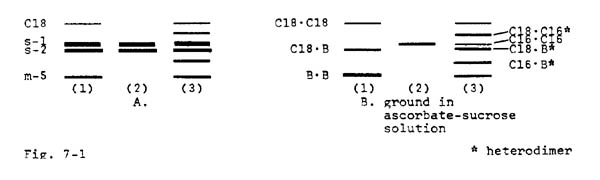
Had photographs of gels been included with these notes (instead of simplified diagrams), the reader would have had difficulty overlooking the coincident variation of the "intermediate" mMDH bands, such as m-1 and m-2 (in Fig. 1-1), with the positions of the "lower" (m-3, m-5, m-7) and "upper" (e.g. C-18) mMDH bands. For example, when the m-5 band is present, there is nearly always a band at the m-2 position. The notable exception to this rule occurs in the H25 line, where C-18 as well as m-5 is present. In this case, there is no mitochondrial band at the m-2 position; however, there is a mitochondrial MDH band migrating to approximately the s-2 position, which is intermediate between C-18 and m-5. One of the clearest illustrations of the formation of heterodimers between the C gene product and the products of the other mitochondrial loci is shown in Figure 7-1. The first pattern is present in the H25 line and pattern two is present in a line kindly provided by Goodman. When these two lines were crossed together the F1 MDH profile in pattern three was observed. In Fig. 7-1B the kernels were ground in the ascorbate-sucrose solution in order to selectively eliminate the soluble bands. An obvious conclusion is that the C locus product can participate in the formation of both allelic and intergenic heterodimers. In short, there is classical genetic evidence supporting the idea that the products of the three identified mitochondrial MDH genes can freely heterodimerize. In addition, evidence from TB-A localization studies strongly support this interpretation.
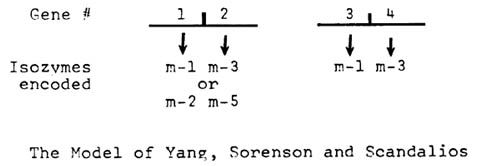
In 1977, N. Yang, J. Sorenson, and J. Scandalios (PNAS 74:310) proposed an alternative genetic model for the mitochondrial MDH genes. They were the first to publish the fact that there is a duplication of the maize mMDH genes. However, in that paper, the authors attempted to explain their m-2 and m-1 bands as products of additional closely linked duplicate genes, as shown below:
This hypothesis is problematical from a mutational point of view since it requires correlated mutation in closely linked genes. When there is a mutation in their Mdh2 gene--leading to an altered electrophoretic mobility of its enzymatic product, a corresponding mutation (leading to a product changed in the same way) must occur in their Mdh1 locus, either subsequently or simultaneously.
The Yang, Sorenson and Scandalios model also requires restrictions on dimer formation. Despite the fact that Yang has demonstrated that the products of two independently assorting mMDH loci--their Mdh2 and Mdh4--form heterodimers (corresponding to band m-4), their similarly related Mdh1 and Mdh3 gene products would not form heterodimers. In addition, their Mdh1 and Mdh2, as well as Mdh3 and Mdh4 gene products, could not heterodimerize. In fact only the products of Mdh2 and Mdh4 would interact on a subunit level. While such heterodimer formation restrictions are not conceptually impossible, our evidence suggests there is free heterodimerization among the products of three independently assorting mitochondrial MDH genes.
The evidence that Yang et al. cite in favor of their model is two-fold: recombinational and biochemical. In the discussion of the paper, the authors report seeing three recombinants (out of 977 tested F2 progeny) between Mdh1 and Mdh2 such that band m-1 was seen with m-5 instead of band m-2 with m-5. Bands m-3 and m-1 were also present from their loci Mdh3 and Mdh4. However, no attempt was made to establish these exceptional progeny as genetic stocks for further analysis. Without such confirmatory evidence, it is extremely difficult to eliminate the possibilities of electrophoretic artifact or, in the absence of contamination markers, fertilization by pollen from other lines.
Biochemical evidence presented by Yang includes the observations that m-1 and m-2 isolated isozymes are very similar in their kinetic properties, while m-3 and m-5 are also similar according to the same criteria. Such evidence is compatible with a heterodimer interpretation for bands m-1 and m-2. Assuming that the mitochondrial gene on chromosome 3 (MdhC) differs in its biochemical properties from the other two mitochondrial loci, then m-1 and m-2--which each have a C-gene subunit--would be expected to be similar to each other, but different from m-3 and m-5.
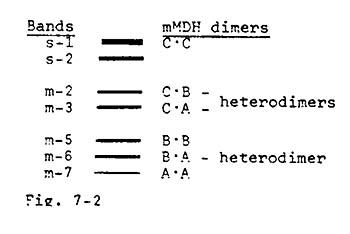
Finally, one can explain on a "heterodimer model" a type of pattern (found in their lines 81 and 6) that was difficult for Yang et al. to interpret on their model. The pattern with our interpretation is illustrated in Figure 7-2. In this case, bands were present at their m-3 and m-5 positions although the m-4 hybrid band, which they expected to see, was missing. On our model, the band at m-3 would not represent a homodimer--rather it would be a heterodimer between two different subunits: the product of the A gene encoding the m-7 homodimer and the C gene product, whose homodimer migrates at the s-1 position.
Kathleen J. Newton
Return to the MNL 53 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}